 肾脏离子转运及利尿剂
肾脏离子转运及利尿剂

《肾脏离子转运及利尿剂》由会员分享,可在线阅读,更多相关《肾脏离子转运及利尿剂(5页珍藏版)》请在装配图网上搜索。
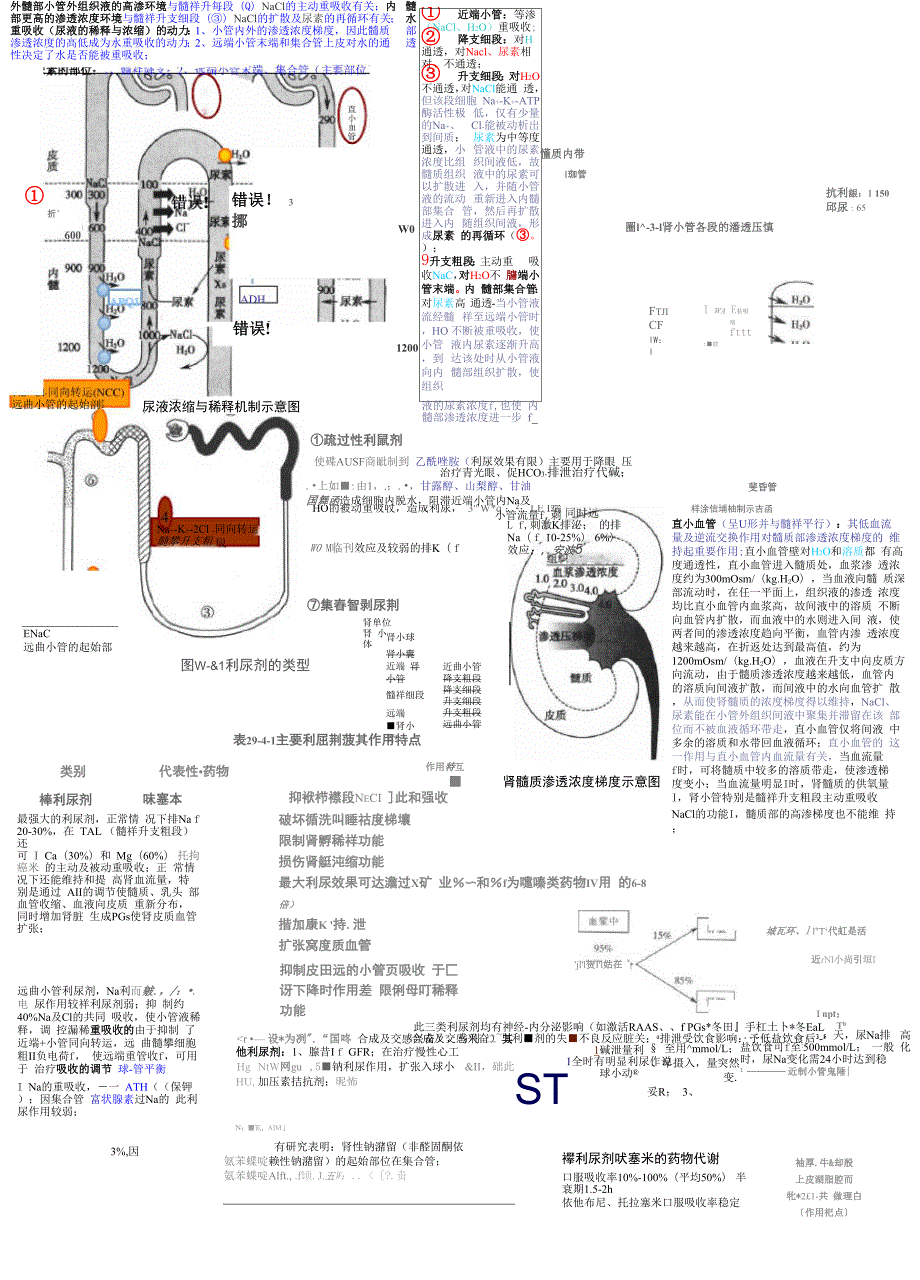
1、外髓部小管外组织液的高渗环境与髓祥升每段(Q)NaCl的主动重吸收有关;内 髓部更高的渗透浓度环境与髓祥升支细段()NaCl的扩散及尿素的再循环有关; 水重吸收(尿液的稀释与浓缩)的动力:1、小管内外的渗透浓度梯度,因此髓质 部渗透浓度的高低成为水重吸收的动力;2、远端小管末端和集合管上皮对水的通 透性决定了水是否能被重吸收;折600远曲小管的起始剖ENaC远曲小管的起始部类别棒利尿剂错误! 3挪错误!尿液浓缩与稀释机制示意图Na+-K+-2Cl -同向转运 髓攀升支粗W01200疏过性利鼠剂 近端小管:等渗 (NaCl、H2O)重吸收; 降支细段:对H 通透,对Nacl、尿素相 对、不通透;
2、 升支细段:对H2O 不通透,对NaCl能通 透,但该段细胞 Na+-K+-ATP酶活性极 低,仅有少量的Na+、 Cl-能被动析出到间质; 尿素为中等度通透,小 管液中的尿素浓度比组 织间液低,故髓质组织 液中的尿素可以扩散进 入,并随小管液的流动 重新进入内髓部集合 管,然后再扩散进入内 随组织间液,形成尿素 的再循环(。);9升支粗段:主动重 吸收NaC,对H2O不 牖端小管末端。内 髓部集合管:对尿素高 通透-当小管液流经髓 祥至远端小管时,HO 不断被重吸收,使小管 液内尿素逐渐升高,到 达该处时从小管液向内 髓部组织扩散,使组织液的尿素浓度f,也使 内髓部渗透浓度进一步 f懂质内带
3、I珈管抗利龈:1 150邱尿:65圈l-3-l肾小管各段的潘透压慎使碟AUSF商眦制到 乙酰唑胺(利尿效果有限)主要用于降眼 压治疗青光眼、促HCO3-排泄治疗代碱;.上如:由1,.;.,甘露醇、山梨醇、甘油国舞函造成细胞内脱水,阻滞近端小管内Na及HO的被动重吸收,造成利尿, 3W*q :2;LE I端小管流量f,刺W0 M临刊效应及较弱的排K( f集舂智剥尿荆图W-&1利尿剂的类型代表性药物味塞本最强大的利尿剂,正常情 况下排Na f 20-30%,在 TAL (髓祥升支粗段)还可 I Ca(30%)和 Mg(60%) 托拘癌米 的主动及被动重吸收;正 常情况下还能维持和提 高肾血流量,特
4、别是通过 AII的调节使髓质、乳头 部血管收缩、血液向皮质 重新分布,同时增加肾脏 生成PGs使肾皮质血管 扩张;远曲小管利尿剂,Na利而皴.,/:.电 尿作用较祥利尿剂弱;抑 制约40%Na及Cl的共同 吸收,使小管液稀释,调 控漏稀重吸收的由于抑制 了近端+小管同向转运,远 曲髓攀细胞粗II负电荷f, 使远端重管收f,可用于 治疗吸收的调节 球-管平衡I Na的重吸收,一 ATH(保钾);因集合管 富状腺素过Na的 此利尿作用较弱;3%,因FtjiCFIW:II WA E粘啜啧 fttt:雇斐昏管肾单位 肾 小 体肾小球肾小囊 近端 肾小管髓祥细段远端肾小管近曲小管 降支粗段 降支细段 升
5、支细段 升支粗段 远曲小管表29-4-1主要利屈荆菠其作用特点L f,刺激K排泌; 的排 Na( f 10-25%) 6%)效应;,.安波5作用特互肾髓质渗透浓度梯度示意图抑袱栉襟段NeCI 此和强收样涂信埔柚制示吉函直小血管(呈U形并与髓祥平行):其低血流 量及逆流交换作用对髓质部渗透浓度梯度的 维持起重要作用;直小血管壁对H2O和溶质都 有高度通透性,直小血管进入髓质处,血浆渗 透浓度约为300mOsm/(kg.H2O),当血液向髓 质深部流动时,在任一平面上,组织液的渗透 浓度均比直小血管内血浆高,故间液中的溶质 不断向血管内扩散,而血液中的水则进入间 液,使两者间的渗透浓度趋向平衡,血
6、管内渗 透浓度越来越高,在折返处达到最高值,约为 1200mOsm/(kg.H2O),血液在升支中向皮质方 向流动,由于髓质渗透浓度越来越低,血管内 的溶质向间液扩散,而间液中的水向血管扩 散,从而使肾髓质的浓度梯度得以维持,NaCl、 尿素能在小管外组织间液中聚集并滞留在该 部位而不被血液循环带走,直小血管仅将间液 中多余的溶质和水带回血液循环;直小血管的 这一作用与直小血管内血流量有关,当血流量f时,可将髓质中较多的溶质带走,使渗透梯 度变小;当血流量明显I时,肾髓质的供氧量I,肾小管特别是髓祥升支粗段主动重吸收破坏循洗叫睡祜度梯壤限制肾孵稀祥功能损伤肾艇沌缩功能最大利尿效果可达澹过X矿
7、业和f为嚏嗪类药物IV用 的6-8倍)揩加康K 持.泄扩张窝度质血管NaCl的功能I,髓质部的高渗梯度也不能维 持;抑制皮田远的小管页吸收 于匚讶下降时作用差 限俐母叮稀释功能城瓦环、l lT代虹是活jll贺Il姑在近rNl小尚引垣II npt;Thr 设*为冽.“国咚 合成及交感兴奋),与利 二其他利尿剂:1、腺昔I f GFR;在治疗慢性心工Hg NtW网gu ,5钠利尿作用,扩张入球小 &II,础此HU, 加压素拮抗剂;昵怖N;瓦,AIM有研究表明:肾性钠潴留(非醛固酮依氨苯蝶啶赖性钠潴留)的起始部位在集合管;氨苯蝶啶Alft., .f顷.J.五W;. ?.责此三类利尿剂均有神经-内分泌
8、影响(如激活RAAS、f PGs*冬田手杠土卜*冬EaL合成及交感兴奋)与利剂的失不良反应脏关;a排泄受饮食影响:予低盐饮食后3-51碱泄量利1全时有明显利尿作说球小动ST 至用mmol/L;、早摄入,量突然变.妥R; 3、,天,尿Na排 高盐饮食可f至:500mmol/L; 一般 化时,尿Na变化需24小时达到稳1近制小管鬼陲|襻利尿剂吠塞米的药物代谢口服吸收率10%-100%(平均50%) 半衰期1.5-2h依他布尼、托拉塞米口服吸收率稳定袖厚.牛&却殷上皮湖脂腔而牝*21-共 做理白作用祀点2、体内AngII的变化管腔心伸)ID7.23.R99.9+2流量、 醛固酮 分泌、数及儿 茶酚胺
9、 释放的 改变密1、AngII低浓度f Na/H2O重吸收Na+-H+、9-41A. RNa+-K+-ATP 酶管腔侧LJa受体激动剂B受体激动剂fNa/H O重吸收Cl-碱基协同交电中性Na-Cl-转运Na+-Cl-协同转运主要是Na+-H+交换、Cl- 基交换蛋白协同作用多巴胺 +基底膜侧M:l图H4-4瞪棒升歪辑段、武1重吸收模型ADH盐皮质激素生电性Na-Cl-转运Na+-K+-ATP 酶将 Na+泵 出基底膜,管腔内Na+ 顺浓度梯度进入细胞内, 与官腔间产生的电位差 驱动Cl-从细胞间隙转 运;特点:1、Na在髓袢粗段的重吸收不是等 渗的,与尿液的浓缩 稀释过程密切相关, 参与构成
10、肾髓质外带 渗透压梯度;2、Na的重吸收与K的重吸收密切相关,互相促进,不再与有机溶质相关联;3、髓祥对钠的 重吸收调节因素发生特点:1、Na在近端小管的重吸收基本上是等渗的(近端小管分布着大量的AQP1,对水的通透性高)2、Na重吸收与其他溶质,如HCO/ GS及AA等的重吸收相互偶联;3、近端小管的各段对Na及各溶质的重吸收不均一(S1段重吸收作用最强,S2段及S3段的重吸收减弱;基础:S2段各段细胞形态不同;S1段(近曲小管,PCT)官腔侧上皮褶皱成多层,形成绒毛,细胞内有大量线粒体;(皮质近直小管)、S3段(髓质近直小管)细胞变平,绒毛及线粒体减少);4、近端小管对Na的重吸收有多个转
11、运通道,分两个阶段:1)主要是Na和中性有机溶质、Na和NaHCO 被动重吸收(主要转运方式:与AA、GS的偶联转运及Na+-H+交换),细胞间隙还存在少量Cl-的被动转运;落近袖小曾FD 5Na+-K+-ATPKNa+-H+、Cl-碱基(有机阴离子)交换1七:L出:小密斑滤过:中于存在球Na-重吸收的调节醛固酮ADHANP与缓激肽近州EflJhJJuKloglofnanjIar cdII田璋小靖除 drIBridlecr图球旁H示意图 Uil律村沏JSS0.他一 ExlrHH|tNnurLiiu MGzsanial或由斑 血7?| oSnsH量明显减少时,肾K的排泄量才会减少;: 1制不明,
12、.JUrArani胞,位%髓小酵町由小首一口 bialial dotim口iLrIWlubulfl膜上出现上皮 性钠通道(ENaC),为醛 固酮的调节点;3、钠的 重吸收伴随着远端肾单位:远 曲小管、连接小 管(又称为后远 端小管或集合 管起始部)、集 合管;特点:肾由钠球K 重吸收效率明管腔侧勺排泌,但是正常上膜细胞核入秋小动脉有广泛摆触;2、表达神经源性NO合成酶(nNOS)及 环氧合酶2 (COX-2);3、可感受流经远端肾小管滤过液中Na+浓 度,通过调节肾素的释放来调节入球小动 脉血管张为,以此来控制肾小球率过滤; 还可通过释放NO抑制球管反馈; 体的成含量与细胞外液量紧密相关, 细
13、胞外液的变化主要体现在血压的变化(血容量的变化)和水肿(细胞间质容量的变田岩伸沼成在近BR胃小管再明收惰况沿近端肾小管各段各种溶质浓度及跨上皮电位情况:主要作用于远端小管及集合管, 通过调节蛋白质的表达发挥生理作用; 作用于H+-ATP酶促进泌H+、也可 通过保Na +排K屣进泌HENaC基底膜侧ENaC上细胞ANP口用细施a2-管腔 侧Na+-K+-ATP排泌:远曲小 靠近,排泌量十质集合管K的;出现K的排泌,但量较小, :渐f;研究显示,远曲小管对K的排泌 官腔内的ci1 -J盐曲应漫妈却远端小晋利集皆管蛔拒的作用酸中毒时,CA活性fCA: S1段有丰富的 CAHCO3-在该处被重 吸收;
14、外髓集合管中 段也有CA的表达, 为远端肾小管重吸 收HCO3-的最重要 部位,其他远端肾单 位不表达CA,不能 有效吸收HCO3-,造 成局部PH下降,可 以促进NH3扩散至 官腔内,与H+结合, 形成NH4+,从而参 与尿的酸化功能; 正常情况下,近端小 管对HCO3-的重吸 收量随滤过的f而 f,到达一定程度即 达饱和,此即重吸收 HCO3-的阈值 (24-27mmo1/L);, 约90%的基底膜侧Na+-H+交换子1)Na+-K+-ATP 酶Na+-K+/NH4+-C1-共转运肾脏在维持酸碱平衡方面的主要功能:1、将滤出的HCO3-重吸收;约90%的HCO3-在近端小管被 重吸收,若近
15、端小管功能异常,HCO3-将从尿中流失,使血HCO3-下降,导致近端肾小管代谢性酸 中毒;2、产生新的HCO3-(肾脏排胺是净酸排泌、产生新HCO3-的主要机制)以补充在缓冲机体 产生H+时消耗的HCO-,同时产生的H+通过可滴定酸和NH4+从肾脏排出;近端小管对H+的分泌及HCO3-的重吸收肾小管分泌的H+需与hco3-结合生成H2O和CO2、与nh3结合生成nh4+、或以可滴定酸的形式才 能被排出体外;肾小管重吸收是以CO2的形式进行的,在CA的作用下,上皮细胞每分泌1个H+ 可使1)Na+个和1HCO3-个重吸收回血液;酸中毒时CA活性增高,泌管腔侧H+C1-HCO3-交换子K+-HCO
16、3 -共转运髓祥升支粗段对H+的分泌及HCO3-的重吸收当尿液pH为4.8时,HPO 2-: H PO -由 4: 1 ::变为41: 99,尿液中的 磷酸盐几乎都已变成了 HPO-,已不能进一步 发挥缓冲作用了;此时 上皮细胞泌NH3进一步i1HPOO,-H在连接小管、皮质集合管参与酸碱代谢的主要 细胞为分散在主细胞之间的插入细胞(ICs);rr/HCQ;交换体及保碱的作强;髓祥升支粗段继续重吸收从近端小管逃逸的hco3-,但因为滤液中的H2OI,最终hco3-浓度仍与近端小管管 腔内一致;同时因超滤液中H2OI,使得CO2游离出来弥散到细胞内,而HCO3-则仍留在小管液中,造成小管 液pH
17、 f,使NH3+H+ NH4+的平衡向左移,于是近端肾小管分泌的NH4+约40-80%就以NH3自由弥散到髓攀升 支粗段并进入肾髓质间质;由于氨的重吸收与水不同步,以后各段平行排列的肾小管之间就产生跨上皮的浓度 差,与肾脏髓质内平行排列的直小血管之间的浓度差一起形成逆流倍增系统,进一步将大量NH3和NH4+聚集 在髓质间质,继而分泌到邻近的髓质集合管内,因此,终尿中排出的铵基本上都来自近端小管合成并被TAL 吸收的氨和铵,而完善的浓缩和逆流倍增系统则是肾脏排铵功IflBHMCl hco3交换* HCO;ATPHATPWiHg远端小管和集合管还可泌H+和泌K+,均 与管腔中Na+交换,分别称为H
18、+-K+交换 和Na+-K+交换,二者之间存在相互抑制;体内pH调节 血液缓冲系统反应最为 迅速,但作用 不持久;肺的 调节效能大, 也很迅速,在 数min内开始 发挥作用, 30min时达最 大,但仅对 CO2有调节 作用;细胞内 液的缓冲能 力虽较强,但 约3-4小时后 才发挥作用; 肾脏的调节 作用发挥更 慢,需数h后 才发挥作用, 3-5天达高 峰,但其作用 强大而持久, 能有效地排 出固定酸,保 留 NaHCO3;中和H +圈l-4-4i-4 K型插入细胞的分泌HCtK机制阳1淅警A,逝端环野上度瞄也明俨11HPtX+irHri+HC0S1Na+-H+HIMC(+HtO其侧膜I fi
19、同蝴 血 ft己成于远曲小管后端和集合管;图A型糖入细胞的祯靠机制K 花 阮 脱氢:谷氨酰胺酶(GT)厂 酸中毒,活性f瓶帅jeA COa*HaOnh 悖ty图1-4书书 肾脏胺的产生及分泌JS海Na.HCO,龄阪蓄积远端小管对H+分泌及HCO3-的重吸收(润细胞/泌H+细胞)e.mafiHco;+trMs HjCDjHiOg管牲腹* H+旺皿eg珀0管趣膜图了- 2近嫌小管和照会曾祕H, 吸攻H8 JS程示童图 丸近藤小管f出细的B 孙竹P反碘HjO+COzNH.E质集合管同时具有泌H+泌 【管腔内为正电位,其泌氢2CA LNH;X5NH:靴;4fl Na+的功能;外髓集hsH+泌H+而不参
20、与K+/Na赖于钠的当尿液pH为4.8时,HPO,2 -已不能进一步 土发挥缓冲作用了,此 瘦运上皮细胞泌NH3进 ;一步中和H+;r+Hco;泌NH3.NH4+的同时保碱回仪HE亍芍-120%NIL75HJIC01虽然肾小管全程的上皮细胞均能合成NH3,但绝大部分还是 合成于近端肾小管的初始段和中段,另外可能还有部分NH4+ 通过Na+-K+(NH4+)-ATP酶从管周被泵入细胞内;细胞内的 NH4+部分通过Na+-NH4+交换排泌到小管液中,部分以NH3直 接弥散到酸性的官腔液中形成NH4+;近端小管分泌的NH4+ 大部分在髓祥升支粗段被重吸收;远曲小管后段和集合管含有两类细胞,即主细胞和
21、闰细胞。主细胞重吸收Na+和水 分泌K+。闰细胞则主要分泌H+。主细胞重吸收Na+主要通过管腔膜上的Na+通道。 管腔内的Na+顺电化学梯度通过管腔膜上的Na+通道进入细胞,然后,由Na+泵泵 至细胞间液而被重吸收。(1)K+的分泌:尿中排出的K+和Na+的来源不同,尿中的 Na+是肾小球滤出后未被肾小管重吸收的多余Na+,而尿中的K+则主要是远球小管闰细胞内的CA催化CO与HO生成HCO,分解成H+与HCO-,H+常 与Na+交换主动分泌,异与K+2形成竞争233幽哗3髓犊升支粗段和内层睁质集合瞥未段F廿转德Ng图2-1-2国出尚肾单位转远的全过程谯示该校NHj浓度为究末尿法度(血潦的西丹数
22、和集合管分泌的。尿中K+的排泄量视K+的摄入量而定。高钾饮食可排出大量的钾, 低钾饮食则尿中排钾量少,即使是无钾饮食时,尿中也会便出钾。但如饮食无钠时, 尿中也无钠排出。说明机体保钠能力明显大于保钾。K+分泌的机制可概括为: 远曲小管后段和集合管的主细胞内的K+浓度明显高于小管液中的K+浓度,K+便 顺浓度梯度从细胞内通过管腔膜上的K+通道进入小管液; 在远曲小管和集合管,主细胞基侧膜上的Na+泵将细胞内的Na+泵至细胞间隙而 被重吸收,小管液中Na+通过主细胞的管腔膜上的Na+通道进入细胞,使管腔内带 负电位(-1040mV)。这种电位梯度也成为K+从细胞内分泌至管腔的动力。这类 Na+泵的
23、活动是生电性的。 进入主细胞的Na+,可刺激基侧膜上的Na+泵,使更多的K+从细胞外液中泵入细 胞内,提高细胞内K+浓度,增加细胞内和小管液之间的K+浓度梯度,从而促进K+ 是顺浓度差分泌。因此,K+的分泌与Na+的重吸收相关联,即所谓的K+-Na+交换作 用;远曲小管和集合管重吸收NaCl、分泌K+和H+的示意图 A:远曲小管初段_B:远曲小管后段和集合管H+的分泌除了近球小管细胞通过Na+-H+交换分泌H+,促进NaHCO3重吸 收外,远曲小管和集合管的闰细胞也可分泌H+。H+的分泌是一个逆电化 学梯度进行的主动转运过程。有人认为管腔膜上有H+泵,能将细胞内的 H+泵入小管腔内。细胞内的C
24、O2和H2O在碳酸酐酶催化作用下生成的H+ 和HCO3-,H+由H+泵泵至小管液,HCO3-则通过基侧膜回到血液中,因而 H+分泌和 HCO-的重吸收与酸碱平衡的调节有关(图B)。闰细胞分泌的 +。H+与HPO2 -结合形成HPO,-,这是可滴定酸;分泌的H+可与上皮细胞分 泌的NH3结合,形成Nh4;。可滴定酸和NH4+都不易透过管腔膜进入细 胞而留在小管液中。因此,它们是尿液酸碱度的决定因素。泌K+与泌H+呈负相关:K+-Na+交换与H+ -Na+交换两者相互竞争抑制,当 酸中毒时H+ f, H+-Na+交换加强,K+-Na+交换I,泌K+ I,可造成血K+ f,反过来用乙酰唑胺一抑制碳酸
25、酐酶(carbonic anhydrase),使 H+ I以纠 正酸中毒,会使H+-Na+交换I,K+-Na+交换f,造成血K+ I。NH3的分泌:远曲小管和集合管的上皮细胞在代谢过程中不断地生成 NH3,这些NH3主要由谷氨酰胺脱氨而来。NH3具有脂溶性,能通过细 胞膜向小管液周围组织间液和小管液自由扩散,扩散量取决于两种液体的 pH值。小管液的pH较低(H+浓度较高),所以NH3较易向小管液中扩散。 分泌的NH3能与小管液中的H+结合并生成NH4+,小管液中NH3浓度因 而下降,于是管腔膜两侧形成了 NH3浓度梯度,此浓度梯度又加速了 NH3 向小管液中扩散。由此可见,NH3的分泌与H+的分泌密切相关;H+分泌 增加促使NH3分泌增多。NH3与H+结合并生成NH4+后,可进一步与小 管液中的强酸盐(如NaCl等)的负离子结合,生成酸性铵盐NH4Cl等) 并随尿排出。强酸盐的正离子(如Na+)则与H+交换而进入肾小管细胞, 然后和细胞内HCO -一起被转运回血。所以 肾小管细胞分泌NH3,不仅 。 匕人,由于铵盐形成而促进了排H+,而且也促进了 NaHCO3的重吸收;
- 温馨提示:
1: 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
2: 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
3.本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
5. 装配图网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
