 病毒的增殖课件
病毒的增殖课件

《病毒的增殖课件》由会员分享,可在线阅读,更多相关《病毒的增殖课件(66页珍藏版)》请在装配图网上搜索。
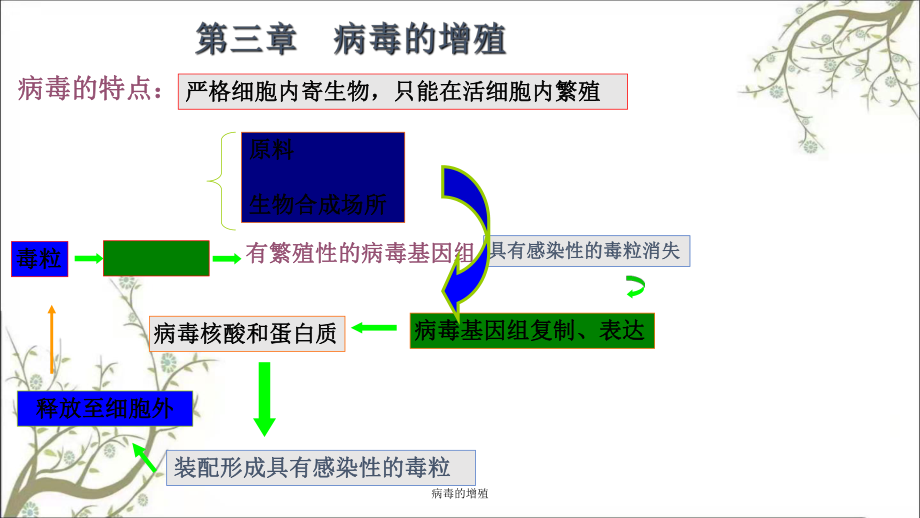
1、病毒的增殖病毒的特点:病毒的特点:严格细胞内寄生物,只能在活细胞内繁殖严格细胞内寄生物,只能在活细胞内繁殖毒粒毒粒有繁殖性的病毒基因组有繁殖性的病毒基因组具有感染性的毒粒消失具有感染性的毒粒消失病毒基因组复制、表达病毒基因组复制、表达病毒核酸和蛋白质病毒核酸和蛋白质装配形成具有感染性的毒粒装配形成具有感染性的毒粒释放至细胞外释放至细胞外原料原料生物合成场所生物合成场所病毒的增殖一、病毒的复制周期一、病毒的复制周期(一)、一步生长曲线(一)、一步生长曲线(二)、病毒侵染寄主细胞的过程(二)、病毒侵染寄主细胞的过程二、病毒的异常复制(非增殖性感染)二、病毒的异常复制(非增殖性感染)病毒的增殖病毒的
2、增殖方式病毒的增殖方式复制复制replication病毒感染敏感宿主细胞后,病毒核酸进入细胞,通过其复制与病毒感染敏感宿主细胞后,病毒核酸进入细胞,通过其复制与表达产生子代病毒基因组和新的蛋白质,然后由这些新合成的表达产生子代病毒基因组和新的蛋白质,然后由这些新合成的病毒组分装配(病毒组分装配(assembly)成子代毒粒,并以一定方式释放到成子代毒粒,并以一定方式释放到细胞外。细胞外。一、病毒的复制周期一、病毒的复制周期复制周期(复制周期(replicative circle)或称复制循环:或称复制循环:自病毒吸附于细胞开始,到子代病毒从感染细胞自病毒吸附于细胞开始,到子代病毒从感染细胞释放
3、到细胞外的病毒复制过程。释放到细胞外的病毒复制过程。病毒的增殖用于病毒复制研究的实验系统用于病毒复制研究的实验系统噬菌体噬菌体细菌培养系统细菌培养系统动物病毒动物病毒动物细胞培养系统动物细胞培养系统植物病毒植物病毒动植物原生质体培养系统动植物原生质体培养系统 易培养、易控制易培养、易控制 噬菌斑、易观察噬菌斑、易观察 生长快,周期短生长快,周期短病毒的增殖一、病毒的复制周期一、病毒的复制周期(一)、一步生长曲线(一)、一步生长曲线(one step growth curve)19391939年,年,Max Delbruck & & Emory Ellis:E. coli / bacteriop
4、age1 1、用噬菌体的稀释液感染高浓度的宿主细胞、用噬菌体的稀释液感染高浓度的宿主细胞2 2、数分钟后,加入抗噬菌体的、数分钟后,加入抗噬菌体的抗血清抗血清3 3、将上述混合物、将上述混合物大量稀释大量稀释4 4、保温培养并定期检测培养物中的噬菌体、保温培养并定期检测培养物中的噬菌体效价效价5 5、以感染时间为横坐标,病毒的感染效价为纵坐标,、以感染时间为横坐标,病毒的感染效价为纵坐标,绘制出病毒特征性的繁殖曲线绘制出病毒特征性的繁殖曲线该实验标志着分子病毒学、分子生物学和分子遗传学该实验标志着分子病毒学、分子生物学和分子遗传学的建立,的建立,Max Delbruck因此荣获因此荣获1969
5、1969年年Nobel Prize病毒的增殖噬菌体的一步生长曲线噬菌体的一步生长曲线吸附期吸附期潜伏期潜伏期裂解期裂解期 (时 间) (时 间)(噬 菌 斑 数)(噬 菌 斑 数)病毒的增殖一、病毒的复制周期一、病毒的复制周期(一)、一步生长曲线(一)、一步生长曲线(one step growth curve)噬菌体复制(繁殖)噬菌体复制(繁殖)的三个阶段:的三个阶段:1 1、吸附期、吸附期游离的噬菌体吸附到宿主细胞游离的噬菌体吸附到宿主细胞2 2、潜伏期、潜伏期从噬菌体吸附到细胞到释放从噬菌体吸附到细胞到释放出新噬菌体的最短时期出新噬菌体的最短时期3 3、裂解期、裂解期随着菌体不断破裂,新噬
6、菌体随着菌体不断破裂,新噬菌体数目增加,直到最高值数目增加,直到最高值病毒的增殖取培养物直接测取培养物直接测定病毒数量定病毒数量将培养物过滤去细将培养物过滤去细胞后测定病毒数量胞后测定病毒数量将细胞裂解后测定病毒的数量将细胞裂解后测定病毒的数量前期可将培养物先离心去上清以前期可将培养物先离心去上清以消除未吸附病毒的影响消除未吸附病毒的影响裂解量:裂解量:每个受染细胞所产生的子代病毒颗粒的平均数目。每个受染细胞所产生的子代病毒颗粒的平均数目。其值等于潜伏期受染细胞的数目除以稳定期受染细胞所释放的全部其值等于潜伏期受染细胞的数目除以稳定期受染细胞所释放的全部子代病毒数目,即等于稳定期病毒效价与潜伏
7、期病毒效价之比。子代病毒数目,即等于稳定期病毒效价与潜伏期病毒效价之比。(一)、一步生长曲线(一)、一步生长曲线(one step growth curve)病毒的增殖隐蔽期(隐蔽期(eclipse period)在潜伏期的前一段,受染细胞内检测不到在潜伏期的前一段,受染细胞内检测不到感染性病毒,后一阶段,感染性病毒在受感染性病毒,后一阶段,感染性病毒在受染细胞内的数量急剧增加。染细胞内的数量急剧增加。自病毒在受染细胞内消失到细胞内出现新自病毒在受染细胞内消失到细胞内出现新的感染性病毒的时间为隐蔽期。的感染性病毒的时间为隐蔽期。隐蔽期病毒在细胞内存在的动力学曲线呈隐蔽期病毒在细胞内存在的动力学
8、曲线呈线性函数线性函数,而非,而非指数关系指数关系,从而证明子代病毒颗粒是由新合成的病毒基因组与蛋白质经装配成从而证明子代病毒颗粒是由新合成的病毒基因组与蛋白质经装配成熟,而不是通过双分裂方式产生的。熟,而不是通过双分裂方式产生的。不同病毒的隐蔽期长短不同不同病毒的隐蔽期长短不同例如,例如,DNA动物病毒的隐蔽期为动物病毒的隐蔽期为5 52020min,RNA动物病毒为动物病毒为2 210 10 h。病毒的增殖(一)、一步生长曲线(一)、一步生长曲线(one step growth curve)是研究病毒复制的一个经典试验,最初是为研究噬菌体的复是研究病毒复制的一个经典试验,最初是为研究噬菌体
9、的复制而建立,以后推广到动物病毒和植物病毒复制研究中。制而建立,以后推广到动物病毒和植物病毒复制研究中。以适量的病毒接种于标准培养的高浓度的敏感细胞,待病毒吸附后,离心除去以适量的病毒接种于标准培养的高浓度的敏感细胞,待病毒吸附后,离心除去未吸附的病毒,或以抗病毒抗血清处理病毒未吸附的病毒,或以抗病毒抗血清处理病毒细胞培养物以建立同步感染,然细胞培养物以建立同步感染,然后继续培养并定时取样测定培养物中的病毒效价,并以感染时间为横坐标,病后继续培养并定时取样测定培养物中的病毒效价,并以感染时间为横坐标,病毒的感染效价为纵坐标,绘制出病毒特征性的繁殖曲线,即毒的感染效价为纵坐标,绘制出病毒特征性的
10、繁殖曲线,即一步生长曲线一步生长曲线。不同病毒的潜伏期长短不同,不同病毒的潜伏期长短不同,噬菌体噬菌体以以minmin计,计,动物病毒动物病毒和和植物病毒植物病毒以以h h或或d d 计。计。潜伏期潜伏期噬菌体噬菌体的裂解量一般为的裂解量一般为几十到上百个几十到上百个,植物病毒和动物病毒植物病毒和动物病毒可可达达数百乃至上万个数百乃至上万个。裂解量裂解量病毒的增殖 吸附吸附adsorption/attachment 侵入侵入penetration/injection 脱壳脱壳uncoating 病毒大分子的合成病毒大分子的合成 病毒基因组的复制病毒基因组的复制与表达与表达 装配装配 assem
11、bly 成熟成熟 maturation 释放释放release一、病毒的复制周期一、病毒的复制周期(二)、病毒侵染寄主细胞的过程(二)、病毒侵染寄主细胞的过程病毒的增殖1 1、吸附、吸附 adsorption/attachment毒毒 粒粒敏感细胞敏感细胞随机碰撞而接触(静电引力或氢键)随机碰撞而接触(静电引力或氢键)可逆吸附,无特异性(可逆吸附,无特异性(非细胞颗粒也可吸附非细胞颗粒也可吸附)病毒表面蛋白与细胞受体的结合病毒表面蛋白与细胞受体的结合特异性,不可逆吸附,特异性,不可逆吸附,启动病毒感染的第一阶段启动病毒感染的第一阶段(二)、病毒侵染寄主细胞的过程(二)、病毒侵染寄主细胞的过程病
12、毒的增殖1 1、吸附、吸附VAP (virus-attachment protein)与细胞受体与细胞受体receptor间的结合力来源于间的结合力来源于空间结构的互补性空间结构的互补性,相互间的,相互间的电电荷荷、氢键氢键、疏水性疏水性相互作用及相互作用及范得华力范得华力。不同种系的细胞具有不同病毒的细胞不同种系的细胞具有不同病毒的细胞受体,受体,病毒受体病毒受体的细胞种系特异性决的细胞种系特异性决定了病毒的宿主范围。定了病毒的宿主范围。病毒的增殖1 1、吸附、吸附分别吸附于大肠杆菌分别吸附于大肠杆菌性毛和菌体上的噬菌体性毛和菌体上的噬菌体病毒的增殖1 1、吸附、吸附 动物宿主细胞及细菌表面
13、病毒受体的存在可通动物宿主细胞及细菌表面病毒受体的存在可通过生化等实验证明;过生化等实验证明; 但但至今仍未发现植物病毒的细胞受体至今仍未发现植物病毒的细胞受体存在。存在。病毒的增殖1 1、吸附、吸附 病毒吸附蛋白病毒吸附蛋白VAP:一般由病毒的衣壳蛋白或包膜上的糖蛋白一般由病毒的衣壳蛋白或包膜上的糖蛋白突起充当。如:突起充当。如: T偶数噬菌体偶数噬菌体噬菌体尾部的噬菌体尾部的尾丝蛋白尾丝蛋白 x174噬菌体噬菌体二十面体衣壳五邻体顶上的纤维突起二十面体衣壳五邻体顶上的纤维突起H蛋白蛋白 无包膜动物病毒如无包膜动物病毒如腺病毒腺病毒二十面体衣壳上的二十面体衣壳上的五邻体纤维五邻体纤维、 多瘤
14、病毒多瘤病毒VPl(衣壳蛋白有衣壳蛋白有VPl、VP2和和VP3) 有包膜动物病毒有包膜动物病毒病毒包膜上的糖蛋白突起,如病毒包膜上的糖蛋白突起,如 副粘病毒副粘病毒HN糖蛋白糖蛋白、流感病毒流感病毒血凝素血凝素( (HA)糖蛋糖蛋白,白, HIVgp120 糖蛋白、糖蛋白、弹状病毒弹状病毒G 糖蛋白,糖蛋白, 疱疹病毒疱疹病毒gpB糖蛋白糖蛋白病毒的增殖1 1、吸附、吸附 在流感病毒包膜上,有两种糖蛋白突起,血凝素在流感病毒包膜上,有两种糖蛋白突起,血凝素( (HA)和神和神经氨酸酶经氨酸酶( (NA) 流感病毒的吸附只与流感病毒的吸附只与HA有关,而与有关,而与NA无关无关 (病毒的感染性
15、只能被抗血凝素的抗血清所中和不能(病毒的感染性只能被抗血凝素的抗血清所中和不能被抗神经氨酸酶的抗血清中和)被抗神经氨酸酶的抗血清中和)流感病毒包膜上血凝素流感病毒包膜上血凝素( (HA)三聚体结构(三亚基)三聚体结构(三亚基)病毒的增殖1 1、吸附、吸附 流感病毒包膜上血凝素流感病毒包膜上血凝素( (HA)分分4 4个区个区1. 1. 细胞受体结合区:细胞受体结合区:流感病毒的流感病毒的VAP与宿主细胞表面糖蛋白受与宿主细胞表面糖蛋白受体的体的唾液酸唾液酸结合,这是结合,这是HA的主要功能其受体结合位点位于的主要功能其受体结合位点位于三聚体的顶端,形似一个裂缝或小口袋。它是病毒包膜糖蛋白三聚体
16、的顶端,形似一个裂缝或小口袋。它是病毒包膜糖蛋白HA与细胞受体相互作用的分子基础。与细胞受体相互作用的分子基础。2. 2. 融合区:融合区: HA蛋白具有促膜融合的活性,但蛋白具有促膜融合的活性,但HA只有经蛋白只有经蛋白酶切割产生酶切割产生HA1和和HA2后才具有融合活性。后才具有融合活性。3. 3. 跨膜区:跨膜区: 在靠近在靠近HA分子的羧基端大约有分子的羧基端大约有2525个个AA残基组成残基组成非极性区即非极性区即HA的跨膜区的跨膜区4. 4. 膜内区:膜内区: HA分子羧基端最后分子羧基端最后1010个个AA在病毒包膜内表面。在病毒包膜内表面。病毒的增殖1 1、吸附、吸附影响病毒吸
17、附的环境因素:影响病毒吸附的环境因素: 噬菌体数量:噬菌体数量: 每一敏感细胞所能吸附相应噬菌体的数量每一敏感细胞所能吸附相应噬菌体的数量m.o.i 温度温度: : 一般来说在一般来说在037范围内温度越高、病毒吸附效率也越高范围内温度越高、病毒吸附效率也越高 温度越高,温度越高,VAP与细胞受体间的碰撞次数愈多,吸附机会越大与细胞受体间的碰撞次数愈多,吸附机会越大 温度越高细胞膜中脂质分子的运动越快,其中蛋白质分子的迁移也温度越高细胞膜中脂质分子的运动越快,其中蛋白质分子的迁移也更加频繁,受体分子更容易与病毒粒子产生多价结合更加频繁,受体分子更容易与病毒粒子产生多价结合 离子:离子:只有在一
18、定的阳离子环境条件下,病毒粒子才能吸附细胞,如只有在一定的阳离子环境条件下,病毒粒子才能吸附细胞,如Na+、K+可使脊髓灰质炎病毒、腺病毒、流感病毒产生最大量的吸附,可使脊髓灰质炎病毒、腺病毒、流感病毒产生最大量的吸附,2价阳离价阳离子子Ca2+、Mg2+则能促进噬菌体吸附则能促进噬菌体吸附 pH:pH的高低能够引起病毒粒子和细胞表面基团电荷的改变(的高低能够引起病毒粒子和细胞表面基团电荷的改变(510510) 其他:其他:某些糖基分子可与细胞受体相互作用,对病毒产生竞争性结合抑某些糖基分子可与细胞受体相互作用,对病毒产生竞争性结合抑制如单糖分子可以抑制人肠道病毒的血凝作用制如单糖分子可以抑制
19、人肠道病毒的血凝作用病毒的增殖2 2、侵入、侵入penetration/injection 侵入侵入(内化内化),一个病毒吸附后几乎立即发生,依赖于,一个病毒吸附后几乎立即发生,依赖于能量的感染步骤。能量的感染步骤。 不同的病毒不同的病毒- -宿主系统的病毒侵入机制不同宿主系统的病毒侵入机制不同 注射式注射式injection 有尾噬菌体有尾噬菌体 细胞内吞细胞内吞endocytosis 动物病毒动物病毒 膜融合膜融合fusion 有包膜的病毒有包膜的病毒 直接侵入直接侵入 部分动物病毒、无包膜病毒、植物病部分动物病毒、无包膜病毒、植物病毒毒病毒的增殖2 2、侵入、侵入penetration/
20、injection(1) (1) 部分病毒粒子可直接侵入部分病毒粒子可直接侵入 如脊髓灰质炎病毒、腺病毒。如脊髓灰质炎病毒、腺病毒。(2) 许多无包膜病毒通过许多无包膜病毒通过VAP与细胞受体结合后,其宿主与细胞受体结合后,其宿主细胞细胞膜的流动性引起病毒衣壳蛋白重排和构型变化膜的流动性引起病毒衣壳蛋白重排和构型变化,细胞膜上,细胞膜上可能有可能有水解酶水解酶,导致病毒粒子释放核酸进入细胞质,病毒,导致病毒粒子释放核酸进入细胞质,病毒衣壳仍留在细胞膜外。实际上将病毒侵入和脱壳融为一体。衣壳仍留在细胞膜外。实际上将病毒侵入和脱壳融为一体。(3) (3) 其他特殊方式其他特殊方式:尚未发现尚未发现
21、植物病毒植物病毒有特异性细胞受体,通有特异性细胞受体,通过过微伤口微伤口或或自然孔口自然孔口、细胞间的、细胞间的胞间连丝胞间连丝,或,或昆虫昆虫的口器的口器侵入细胞侵入细胞。 许多病毒并非仅有一种侵入方式,可能几种侵入方式并存许多病毒并非仅有一种侵入方式,可能几种侵入方式并存直接侵入直接侵入病毒的增殖2 2、侵入、侵入有尾噬菌体有尾噬菌体注射方式将噬菌体核酸注入细胞注射方式将噬菌体核酸注入细胞通过尾部刺突固着于细胞;通过尾部刺突固着于细胞;尾部的酶水解细胞壁的肽聚糖,使细胞壁产生小孔;尾部的酶水解细胞壁的肽聚糖,使细胞壁产生小孔;尾鞘收缩,核酸通过中空的尾管压入胞内,蛋白质外壳留在胞外;尾鞘收
22、缩,核酸通过中空的尾管压入胞内,蛋白质外壳留在胞外;吸附尾钉固着尾鞘收缩尾管穿入DNA注入如果大量噬菌体在短时间内吸附于同一细胞上,使细胞壁产如果大量噬菌体在短时间内吸附于同一细胞上,使细胞壁产生许多小孔,也可引起细胞立即裂解,但噬菌体并未增殖,生许多小孔,也可引起细胞立即裂解,但噬菌体并未增殖,这种现象称为这种现象称为自外裂解自外裂解(lysis from without)。)。病毒的增殖2 2、侵入、侵入动物病毒动物病毒直接穿过细胞膜的移位方式直接穿过细胞膜的移位方式细胞的内吞功能细胞的内吞功能毒粒包膜与细胞质毒粒包膜与细胞质 膜的融合膜的融合病毒的增殖2 2、侵入、侵入植物病毒植物病毒通
23、过人为或自然的机械损伤所形成的通过人为或自然的机械损伤所形成的微伤口微伤口进入细胞;进入细胞;或者靠携带有病毒的媒介,主要靠是有或者靠携带有病毒的媒介,主要靠是有吮吸式口器的昆虫吮吸式口器的昆虫取取食将病毒带入细胞。食将病毒带入细胞。植物病毒一旦进入细胞后,增殖产生的子代病毒或病毒核酸植物病毒一旦进入细胞后,增殖产生的子代病毒或病毒核酸可通过病毒编码的可通过病毒编码的运动蛋白运动蛋白(movement proteinmovement protein)与与胞间连胞间连丝的相互作用丝的相互作用从受染细胞进入邻近细胞。从受染细胞进入邻近细胞。病毒的增殖3 3、脱壳、脱壳 uncoating 病毒侵入
24、后,病毒的包膜和病毒侵入后,病毒的包膜和/ / 或壳体除去而释放出病或壳体除去而释放出病毒核酸的过程。毒核酸的过程。 脱壳是病毒基因组进行功能表达所必需的感染事件。脱壳是病毒基因组进行功能表达所必需的感染事件。T-偶数噬菌体脱壳与侵入是一起发生的偶数噬菌体脱壳与侵入是一起发生的动物病毒存在不同的结构类型和不同动物病毒存在不同的结构类型和不同的侵入方式,其脱壳过程也较复杂。的侵入方式,其脱壳过程也较复杂。病毒的毒粒消失,失去原有的感染性,进入潜隐期。病毒的毒粒消失,失去原有的感染性,进入潜隐期。病毒的增殖3 3、脱壳、脱壳 uncoating病毒的增殖遗传的中心法则遗传的中心法则 转录 翻译复制
25、 DNA mRNA Protein 逆转录(半保留不连续复制)(半保留不连续复制)4、病毒大分子的合成、病毒大分子的合成 ( (replication)全保留复制模式:全保留复制模式:多数多数dsRNA病毒病毒病毒的增殖4、病毒大分子的合成、病毒大分子的合成病毒基因组的复制与表达存在着强烈的时序性病毒基因组的复制与表达存在着强烈的时序性病毒基因组进入胞内病毒基因组进入胞内宿主细胞的代谢发生改变宿主细胞的代谢发生改变病毒利用宿主的生物合成机构和场所,使病毒核病毒利用宿主的生物合成机构和场所,使病毒核酸表达和复制,产生大量的病毒酸表达和复制,产生大量的病毒蛋白质蛋白质和和核酸核酸。病毒的增殖早期蛋
26、白:中期早期蛋白:中期mRNA聚合酶、更改蛋白质等聚合酶、更改蛋白质等中期蛋白:中期蛋白:DNA聚合酶、晚期聚合酶、晚期mRNA聚合酶等聚合酶等晚期蛋白:晚期蛋白:头部蛋白、尾部蛋白、装配蛋白、溶菌酶等头部蛋白、尾部蛋白、装配蛋白、溶菌酶等病毒的增殖 dsDNADNA肝炎病毒肝炎病毒具有具有RNA中间体中间体+ +mRNA +DNA细小病毒科细小病毒科 +RNA逆转录病毒逆转录病毒 dsDNA 腺病毒、疱疹病毒腺病毒、疱疹病毒 +RNA冠状病毒科冠状病毒科蛋白质蛋白质 dsRNA呼肠孤病毒科呼肠孤病毒科 -RNA正粘病毒科正粘病毒科病毒的基因组类型及复制表达途径病毒的基因组类型及复制表达途径d
27、sDNA dsDNA -ssDNA +RNAdsDNA发生在成熟过程中的病毒粒子内发生在成熟过程中的病毒粒子内病毒的增殖Transcription of virus genomes病毒的增殖逆病毒复制:逆病毒复制:mRNA潜伏潜伏杂种双链杂种双链病毒的增殖逆病毒复制过程逆病毒复制过程1:、进入细胞进入细胞;、反转录反转录 在反转录酶的作用下以病毒在反转录酶的作用下以病毒RNA为模为模板合成板合成DNA,然后再形成线形然后再形成线形dsDNA;、整整 合合 DNA拷贝整合到宿主基因组中;拷贝整合到宿主基因组中;、转转 录录 病毒病毒DNA转录形成病毒转录形成病毒mRNA和子代病和子代病毒毒RNA
28、;、壳体化壳体化 在细胞质中产生核衣壳;在细胞质中产生核衣壳;、出出 芽芽 粒子在细胞质膜处以出芽方式放出粒子在细胞质膜处以出芽方式放出. . HIV replication 3HIV2病毒的增殖主要是一种主要是一种DNA聚合酶,表现出三种酶活性:聚合酶,表现出三种酶活性:以以RNA为模板合成为模板合成DNA(反转录反转录) );以以DNA为模板合成为模板合成DNA;RNA酶酶H活性活性( (降解降解RNA:DNA杂交链中的杂交链中的RNA链链) ) 反转录酶在进行反转录酶在进行DNA合成时需要的合成时需要的引物引物是特异的细胞内是特异的细胞内tRNA 用作引物的用作引物的tRNA的类型依病毒
29、种类而不同,是从以前的类型依病毒种类而不同,是从以前的宿主细胞带入病毒粒子的。的宿主细胞带入病毒粒子的。如:如:Rous sarcoma(肉瘤肉瘤) virus使用色氨酸使用色氨酸tRNA。 逆转录酶逆转录酶 reverse transcriptase病毒的增殖从逆转录从逆转录v形成形成dsDNA的步骤的步骤1 1病毒的增殖从逆转录从逆转录v形成形成dsDNA的步骤的步骤2 2病毒的增殖逆逆转转录录病病毒毒的的复复制制+RNA病毒的增殖DNA肿瘤病毒引起细胞转化的模式图肿瘤病毒引起细胞转化的模式图dsDNA如如SV40病毒的全部或部分病毒的全部或部分DNA整合到宿主细胞整合到宿主细胞DNA中;
30、中;编码转化信息的病毒基因被转录并被剪切为病毒编码转化信息的病毒基因被转录并被剪切为病毒mRNA分子,然后转运到胞浆被翻译成分子,然后转运到胞浆被翻译成转化蛋白或转化蛋白或T抗原抗原。T抗原能使得宿主细胞转变为癌细胞抗原能使得宿主细胞转变为癌细胞病毒的增殖-RNA病毒增殖病毒增殖如:弹状病毒如:弹状病毒病毒的增殖单纯疱疹病毒复制单纯疱疹病毒复制dsDNA病毒的增殖5、病毒的装配、成熟与释放、病毒的装配、成熟与释放装配装配( (assembly) ):亦称亦称形态发生形态发生(morphogenesis)新合成的毒粒结构组分组装成完整的病毒颗粒新合成的毒粒结构组分组装成完整的病毒颗粒释放释放re
31、lease成熟的子代病毒颗粒然后依一定途径释放到细胞外成熟的子代病毒颗粒然后依一定途径释放到细胞外病毒的释放标志病毒复制周期结束病毒的释放标志病毒复制周期结束成熟(成熟(maturation)涉及病毒粒子的结构变化(蛋白构型),抗原差异涉及病毒粒子的结构变化(蛋白构型),抗原差异病毒的增殖T4噬菌体的装配是一个极噬菌体的装配是一个极为复杂的自我装配的过程为复杂的自我装配的过程病毒的增殖大多数噬菌体都是以裂解细胞方式释放大多数噬菌体都是以裂解细胞方式释放病毒的增殖丝杆噬菌体(如丝杆噬菌体(如M13M13或或fdfd)不杀死细胞,子代毒粒以分泌方式不杀死细胞,子代毒粒以分泌方式不断从受染细胞中释放
32、,并不断从受染细胞中释放,并同时完成毒粒的组装同时完成毒粒的组装。5、病毒的装配、成熟与释放、病毒的装配、成熟与释放病毒的增殖有有包膜病毒的装配与释放包膜病毒的装配与释放有时也是同时完成的有时也是同时完成的病毒的增殖有包膜的动物病毒是在从宿主细胞核芽出或细胞有包膜的动物病毒是在从宿主细胞核芽出或细胞质膜芽出的过程中裹上包膜而形成包膜病毒质膜芽出的过程中裹上包膜而形成包膜病毒病毒的增殖TMV的装配的装配:RNA穿过螺旋的中心孔并在生长端形成一个可移动的环。穿过螺旋的中心孔并在生长端形成一个可移动的环。病毒的增殖吸附吸附侵入侵入早期:病毒特异性酶的合成早期:病毒特异性酶的合成病毒核酸复制病毒核酸复
33、制病毒结构蛋白质合成病毒结构蛋白质合成装配装配释放释放病毒大分子合成病毒大分子合成脱壳脱壳成熟成熟病毒的增殖裸病毒生活周期裸病毒生活周期病毒的增殖二二 病毒的非增殖性感染病毒的非增殖性感染增殖性感染(增殖性感染(productive infection)感染发生在病毒能在其内完成复制循环的允许细胞内,感染发生在病毒能在其内完成复制循环的允许细胞内,并以并以有感染性病毒子代产生有感染性病毒子代产生为特征。为特征。非增殖性感染(非增殖性感染(non productive infection)感染由于病毒、或是细胞的原因,致使病毒的复制在病毒进感染由于病毒、或是细胞的原因,致使病毒的复制在病毒进入敏
34、感细胞后的某一阶段受阻,结果导致病毒感染的不完全入敏感细胞后的某一阶段受阻,结果导致病毒感染的不完全循环循环。在此过程中,由于病毒与细胞的相互作用,虽然亦可在此过程中,由于病毒与细胞的相互作用,虽然亦可能导致细胞发生某些变化,甚至产生细胞病变,但在受染细能导致细胞发生某些变化,甚至产生细胞病变,但在受染细胞内,胞内,不产生有感染性的病毒子代。不产生有感染性的病毒子代。病毒的增殖1 1、流产感染、流产感染(abortive infection) 依赖于依赖于细胞细胞的流产感染:的流产感染:如病毒感染的细胞是病毒不能复如病毒感染的细胞是病毒不能复制的非允许细胞制的非允许细胞( (nonpermis
35、sive cell) ),将导致流产感染将导致流产感染 依赖于依赖于病毒病毒的流产感染的流产感染: :由基因组不完整的缺损病毒由基因组不完整的缺损病毒(defective virus)引起。这类病毒因一个或多个病毒复制必引起。这类病毒因一个或多个病毒复制必需基因有缺损,丧失了其功能,所以它们无论是感染允许需基因有缺损,丧失了其功能,所以它们无论是感染允许细胞还是非允许细胞,都不能完成复制循环。细胞还是非允许细胞,都不能完成复制循环。病毒的非增殖性感染病毒的非增殖性感染病毒的增殖2 2、限制性感染、限制性感染(restrictive infection)因细胞的瞬时允许性产生,导致因细胞的瞬时允
36、许性产生,导致: : (1) (1) 病毒持续存在于受染细胞内不能复制,直到细胞成为病毒持续存在于受染细胞内不能复制,直到细胞成为 允许性细胞,病毒才能繁殖;允许性细胞,病毒才能繁殖; (2) (2) 一个细胞群体中仅有少数细胞产生病毒子代。一个细胞群体中仅有少数细胞产生病毒子代。 3、潜伏感染(、潜伏感染(latent infection)在受染细胞内有病毒基因组持续存在,但并无感染性病毒颗粒在受染细胞内有病毒基因组持续存在,但并无感染性病毒颗粒产生,受染细胞也不会被破坏。产生,受染细胞也不会被破坏。潜伏感染的另一个极端情况是由于病毒基因的功能表达导致宿潜伏感染的另一个极端情况是由于病毒基因
37、的功能表达导致宿主基因表达的改变主基因表达的改变恶性细胞。恶性细胞。病毒的增殖温和噬菌体的溶源性反应温和噬菌体的溶源性反应烈性噬菌体(烈性噬菌体(virulent phage)感染宿主细胞后能在细胞内正常复制并最终杀死细胞,感染宿主细胞后能在细胞内正常复制并最终杀死细胞,形成裂解循环(形成裂解循环(lytic cycle)。)。温和噬菌体或称溶源性噬菌体(温和噬菌体或称溶源性噬菌体(lysogenic phage)感染宿主细胞后不能完成复制循环,噬菌体基因组感染宿主细胞后不能完成复制循环,噬菌体基因组长期存在于宿主细胞内,没有成熟噬菌体产生。长期存在于宿主细胞内,没有成熟噬菌体产生。这一现象称
38、做溶源性(这一现象称做溶源性(lysogeny)现象现象在大多数情况下,温和噬菌体的基因组都整合于宿主染色体中在大多数情况下,温和噬菌体的基因组都整合于宿主染色体中(如(如噬菌体)噬菌体),亦有少数是以质粒形成存在,亦有少数是以质粒形成存在( (如如P1噬菌体噬菌体) )。病毒的增殖温和噬菌体的溶源性反应温和噬菌体的溶源性反应整合于细菌染色体或以质粒形成存在的温和噬菌体整合于细菌染色体或以质粒形成存在的温和噬菌体基因组称做原噬菌体(基因组称做原噬菌体(prophage)在原噬菌体阶段,噬菌体的复制被抑制,宿主细胞在原噬菌体阶段,噬菌体的复制被抑制,宿主细胞正常地生长繁殖,而噬菌体基因组与宿主细
39、菌染色正常地生长繁殖,而噬菌体基因组与宿主细菌染色体同步复制,并随细胞分裂而传递给子代细胞。体同步复制,并随细胞分裂而传递给子代细胞。细胞中含有以原噬菌体状态存在的温和噬菌体基细胞中含有以原噬菌体状态存在的温和噬菌体基因组的细菌称溶源性细菌(因组的细菌称溶源性细菌(lysogenic bacteria)溶源性细菌经自发裂解或诱发裂解,进入裂解循环溶源性细菌经自发裂解或诱发裂解,进入裂解循环病毒的增殖溶源性感染对细胞的影响溶源性感染对细胞的影响被温和噬菌体感染后形成的溶源性细菌具有被温和噬菌体感染后形成的溶源性细菌具有“免疫性免疫性”,即其它同类噬菌体,即其它同类噬菌体虽然可以再次感染该细胞,但
40、不能增殖,也不能导致溶源性细菌裂解。虽然可以再次感染该细胞,但不能增殖,也不能导致溶源性细菌裂解。免疫性是由原噬菌体产生的阻遏蛋白的可扩散性质所决定的。免疫性是由原噬菌体产生的阻遏蛋白的可扩散性质所决定的。溶源性细菌有时还能获得一些新的生理特性,例如白喉杆菌只有在含有特定溶源性细菌有时还能获得一些新的生理特性,例如白喉杆菌只有在含有特定类型的原噬菌体时才能产生白喉毒素,引起被感染机体发病。类型的原噬菌体时才能产生白喉毒素,引起被感染机体发病。原噬菌体引起的溶源性细菌除免疫性外的其他表形改变,包括原噬菌体引起的溶源性细菌除免疫性外的其他表形改变,包括溶源菌溶源菌细胞表面性质细胞表面性质的改变和的
41、改变和致病性致病性转变被称为溶源转变转变被称为溶源转变( (lysogenic conversion)。溶源菌中的温和噬菌体基因组通常不影响细胞的繁殖功能,溶源菌中的温和噬菌体基因组通常不影响细胞的繁殖功能,但它们可能引起其他的细胞变化。但它们可能引起其他的细胞变化。(1) (1) 免疫性免疫性(2) (2) 溶源转变溶源转变病毒的增殖噬菌体的的溶源性反应噬菌体的的溶源性反应进入宿主后线状基因组进入宿主后线状基因组依靠粘性末端环化依靠粘性末端环化病毒的增殖噬菌体的基因组噬菌体的基因组dsDNA,总长度为,总长度为4850248502bp。DNA分子的两端、各有一条由分子的两端、各有一条由121
42、2个核苷酸组成的彼此个核苷酸组成的彼此完全互补的完全互补的55单链突出序列,即粘性末端。单链突出序列,即粘性末端。控制控制噬菌体复制转录的基因:噬菌体复制转录的基因: 编码头部与包装蛋白的基因编码头部与包装蛋白的基因( (A-F) 尾部基因尾部基因编码尾部蛋白编码尾部蛋白( (z-J) 重组基因重组基因int, xis, exo, red, gam等基因与识别位等基因与识别位att ,其功能是行使整合、重组和切割其功能是行使整合、重组和切割 正调控基因正调控基因N和和Q,N为抗转录终止基因,为抗转录终止基因,Q为抗为抗终止子基因,终止子基因,Q蛋白为晚期正调控因子蛋白为晚期正调控因子 负调控基
43、因负调控基因CI、CII、CIII和和cro DNA合成基因合成基因O、P基因基因 裂解基因裂解基因S、R基因基因病毒的增殖噬菌体的的溶源性反应噬菌体的的溶源性反应基因组的表达与复制存在着强烈的时序性基因组的表达与复制存在着强烈的时序性病毒的增殖噬菌体的基因组噬菌体的基因组噬菌体的基因组:噬菌体的基因组: (4 4个基因簇)个基因簇) 调节区调节区( (regulation) 重组有关的区域重组有关的区域( (recombination) 复制区复制区( (replication) 结构基因区结构基因区( (包括头、尾、装配和裂解有关的基因包括头、尾、装配和裂解有关的基因) ) 基因组有三个调
44、节基因基因组有三个调节基因N、Cro和和QN蛋白抗终止子蛋白抗终止子t1,tR1和和tR2Q蛋白抗终止子蛋白抗终止子tR4cro蛋白对蛋白对CI和和Cro基因的转录起负调控作用基因的转录起负调控作用 病毒的增殖噬菌体的的溶源性反应噬菌体的的溶源性反应溶源化有关的有三个基因溶源化有关的有三个基因CI、C和和C C蛋白使蛋白使C蛋白稳定;蛋白稳定; C/C蛋白能使蛋白能使CI基因启始转录;基因启始转录; CI蛋白是蛋白是阻遏物阻遏物,能使与复制成熟和裂解等有关,能使与复制成熟和裂解等有关的基因都不转录,使的基因都不转录,使噬菌体处于溶源状态。噬菌体处于溶源状态。 噬菌体的噬菌体的溶源途径溶源途径C
45、I蛋白的调控作用:蛋白的调控作用:CI蛋白也称为蛋白也称为阻遏蛋白,是一种阻遏蛋白,是一种酸性蛋白,酸性蛋白,PR操纵子最终被操纵子最终被CI阻遏物所抑制,导致阻遏物所抑制,导致 C蛋白的合成停止,在处于原噬菌体状态时其蛋白的合成停止,在处于原噬菌体状态时其CI蛋白的合成一直维持低速率状态。蛋白的合成一直维持低速率状态。病毒的增殖噬菌体的噬菌体的溶菌途径溶菌途径:v噬菌体究竟是进入溶原途径,还是溶菌途径,噬菌体究竟是进入溶原途径,还是溶菌途径,主要主要受受cro和和CI蛋白表达水平的调控蛋白表达水平的调控,而,而CI蛋白的合成又蛋白的合成又同同C的活性有关。的活性有关。 cro蛋白作为一种阻遏
46、蛋白、抑制溶原状态的建立蛋白作为一种阻遏蛋白、抑制溶原状态的建立 当环境因素及宿主的遗传背景或目前尚不清楚的因当环境因素及宿主的遗传背景或目前尚不清楚的因素使素使C蛋白比蛋白比Cro蛋白相对浓度低时,蛋白相对浓度低时,Cro蛋白蛋白阻阻止止CI转录,不产生转录,不产生阻遏物,使阻遏物,使的其他基因可以转的其他基因可以转录,当产生的录,当产生的Q蛋白的量达到使蛋白的量达到使PR2转录完成时、进转录完成时、进行裂解循环途径。行裂解循环途径。 病毒的增殖噬菌体的的溶源性反应噬菌体的的溶源性反应早期基因表达,产生早期基因表达,产生gpcII,gpcro及及gpcIII,后者可防止宿主的裂解后者可防止宿
47、主的裂解酶对酶对gpcII的降解。的降解。gpcII的积累促使阻遏蛋白的积累促使阻遏蛋白CI的表达的表达阻遏蛋白阻遏蛋白CI的积累导致噬菌体基因的积累导致噬菌体基因转录的终止,形成原噬菌体。转录的终止,形成原噬菌体。早期表达的早期表达的gpcro与与cI的竞争最终的竞争最终确定确定噬菌体是进入溶源状态,噬菌体是进入溶源状态,还是进入裂解循环。还是进入裂解循环。(gpcro表达得早,但与基因组表达得早,但与基因组的结合能力较的结合能力较cI弱)弱)阻遏蛋白cI的同样可以抑制其它新侵入的噬菌体的表达,从而使溶源性细菌具有“免疫性”阻遏蛋白阻遏蛋白CI在一般情况下通过自身的转录激活保持低水平的表达,
48、但有时种种在一般情况下通过自身的转录激活保持低水平的表达,但有时种种原因转录水平下降,会偶尔导致溶源性噬菌体进入裂解循环原因转录水平下降,会偶尔导致溶源性噬菌体进入裂解循环(1010- -5 510 10 - - 2 2)。病毒的增殖噬菌体的溶源性反应噬菌体的溶源性反应通过特定位点整合通过特定位点整合/ /切离细菌染色体切离细菌染色体病毒的增殖噬菌体的的溶源性反应噬菌体的的溶源性反应外界因素如紫外线可外界因素如紫外线可引起宿主染色体的引起宿主染色体的破坏,宿主产生应急破坏,宿主产生应急反应合成具有反应合成具有DNA重重组活性的组活性的RecA蛋白,蛋白,导致导致CI的被降解,的被降解,噬菌体进入裂解循环噬菌体进入裂解循环诱发裂解是检查是否存在诱发裂解是检查是否存在 溶原性细菌的有效方法溶原性细菌的有效方法溶源菌菌落溶源菌菌落 溶源菌及其特殊噬菌斑的形态溶源菌及其特殊噬菌斑的形态(少量的溶源菌与大量敏感菌混合少量的溶源菌与大量敏感菌混合 )病毒的增殖本章要点本章要点病毒的一步生长曲线及其作用;病毒的一步生长曲线及其作用;病毒增殖的一般过程,不同种类病毒增殖过程病毒增殖的一般过程,不同种类病毒增殖过程特点;特点;病毒的基因组类型及复制表达途径;病毒的基因组类型及复制表达途径;病毒的非增殖性感染类型、特点及有关概念。病毒的非增殖性感染类型、特点及有关概念。病毒的增殖
- 温馨提示:
1: 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
2: 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
3.本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
5. 装配图网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
