 2012信号转导
2012信号转导

《2012信号转导》由会员分享,可在线阅读,更多相关《2012信号转导(78页珍藏版)》请在装配图网上搜索。
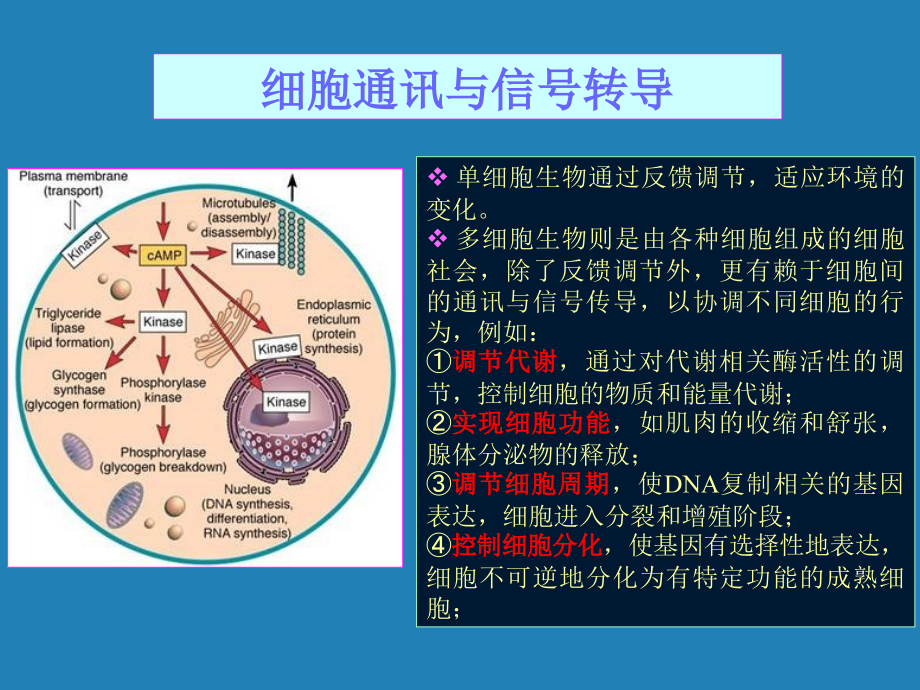
1、细胞通讯与细胞通讯与信号转导信号转导v单细胞生物通过反馈调节,适应环境的变化。v多细胞生物则是由各种细胞组成的细胞社会,除了反馈调节外,更有赖于细胞间的通讯与信号传导,以协调不同细胞的行为,例如:调调节节代代谢谢,通过对代谢相关酶活性的调节,控制细胞的物质和能量代谢;实实现现细细胞胞功功能能,如肌肉的收缩和舒张,腺体分泌物的释放;调调节节细细胞胞周周期期,使DNA复制相关的基因表达,细胞进入分裂和增殖阶段;控控制制细细胞胞分分化化,使基因有选择性地表达,细胞不可逆地分化为有特定功能的成熟细胞;v 信信号号转转导导(signal transduction):细细胞胞外外信信号号分分子子与与胞胞膜
2、膜或或胞胞内内受受体体相相互互作作用用,通通过过信信号号转转换换把把细细胞胞外外信信号号转转变变为为细细胞胞内内信信号号,诱诱发发细细胞胞对对外外界界信信号号作作出出相相应应的的反反应应,这一这一过程称为信号转导过程称为信号转导。一、相关概念:一、相关概念:v 细胞通讯(细胞通讯(cell communication):):指一个细胞发出指一个细胞发出的信息通过介质传递到另一个细胞产生相应反应的过程。的信息通过介质传递到另一个细胞产生相应反应的过程。v受体(受体(receptor):):是一种具有特定功能的蛋白质,存在于细胞是一种具有特定功能的蛋白质,存在于细胞膜上或细胞核内,它能接受外界信号
3、并将这一信号转化为细胞内一系列膜上或细胞核内,它能接受外界信号并将这一信号转化为细胞内一系列生物化学反应,对细胞的结构或功能产生影响。生物化学反应,对细胞的结构或功能产生影响。v配体配体(ligand):受体所接受的外界信号统称为受体所接受的外界信号统称为配体配体,包括激素、,包括激素、神经递质、化学介质、细胞因子、生长因子及其他细胞外信号等,这些神经递质、化学介质、细胞因子、生长因子及其他细胞外信号等,这些细胞外信号又可称为细胞外信号又可称为第一信使(第一信使(first messenger)。v第二信使(第二信使(second messenger):受体将外界信号分子所携带受体将外界信号分
4、子所携带的信号转变为细胞内信号分子,称为信号转导途径中的第二信使,例如的信号转变为细胞内信号分子,称为信号转导途径中的第二信使,例如cAMP、cGMP、Ca 2+等。等。v 胞内信号的转导途径,最终转化为细胞的各种复杂的生物胞内信号的转导途径,最终转化为细胞的各种复杂的生物学效应。学效应。细胞信号转导相关概念:细胞信号转导相关概念:信号信号细胞细胞反应反应增殖增殖分化分化代谢代谢功能功能应激应激凋亡凋亡 or or lostdisease亲亲水水性性信信号号分分子子亲脂性信号分子亲脂性信号分子物理信号(光、物理信号(光、热、电流)热、电流)配体从溶解性来看又可分为配体从溶解性来看又可分为脂溶性
5、和水溶性脂溶性和水溶性两类。两类。脂脂溶溶性性信信号号分分子子,如如甾甾类类激激素素和和甲甲状状腺腺素素,可可直直接接穿穿膜膜进进入入靶靶细胞,与胞内受体结合形成激素细胞,与胞内受体结合形成激素-受体复合物,调节基因表达。受体复合物,调节基因表达。水水水水溶溶溶溶性性性性信信信信号号号号分分分分子子子子,如如神神经经递递质质、细细胞胞因因子子和和水水溶溶性性激激素素,不不能能穿穿过过靶靶细细胞胞膜膜,只只能能与与膜膜受受体体结结合合,经经信信号号转转换换机机制制,通通过过胞胞内内信信使使(如如cAMP)或或激激活活膜膜受受体体的的激激酶酶活活性性(如如受受体体酪酪氨氨酸酸激激酶酶),引引起起细
6、细胞胞的的应应答答反反应应。所所以以这这类类信信号号分分子子又又称称为为第第一一信信使使(primary messenger),而而cAMP这这样样的的胞胞内内信信号号分分子子被被称称为为第第二信使二信使(secondary messenger)。)。目目前前公公认认的的第第二二信信使使有有cAMP、cGMP、三三磷磷酸酸肌肌醇醇(IP3)和和二二酰酰基基甘甘油油(DG),Ca2+被被称称为为第第三三信信使使是是因因为为其其释释放放有有赖赖于于第第二二信使。第二信使的作用是对胞外信号起转换和放大的作用。信使。第二信使的作用是对胞外信号起转换和放大的作用。大多数脂溶性配体大多数脂溶性配体大多数脂
7、溶性配体大多数脂溶性配体可以穿过细胞膜等膜性结构,如甲状腺素、可以穿过细胞膜等膜性结构,如甲状腺素、甾类激素和视黄醛等,它们在合成之后在血液中由载体蛋白协助甾类激素和视黄醛等,它们在合成之后在血液中由载体蛋白协助运输,到达预定位置后与载体蛋白分离,穿过膜性结构。运输,到达预定位置后与载体蛋白分离,穿过膜性结构。脂溶性脂溶性配体配体穿过膜性结构穿过膜性结构有些与胞浆内特异有些与胞浆内特异受体结合形成受体受体结合形成受体-配体复合物配体复合物有些与核内特异有些与核内特异受体结合形成受受体结合形成受体体-配体复合物配体复合物作用于作用于DNADNA分子的特定分子的特定序列,调节某些基因序列,调节某些
8、基因的转录活动的转录活动vCell must respond appropriately to external stimuli to survive.vCells respond to stimuli via cell signalingvSignal transduction pathways consist of a series of steps第一节第一节 受受 体体v概念概念:受体(:受体(receptor)是一种蛋白质,存在于细是一种蛋白质,存在于细胞膜上或细胞核内,它能接受外界信号并将这一信号转胞膜上或细胞核内,它能接受外界信号并将这一信号转化为细胞内的一系列生物化学反应,从而
9、对细胞的结构化为细胞内的一系列生物化学反应,从而对细胞的结构或功能产生影响。或功能产生影响。v 受体的类型受体的类型:根据靶细胞上受体存在部位,受体分根据靶细胞上受体存在部位,受体分为为细胞表面受体(细胞表面受体(cell surface receptor)和和细胞内受体细胞内受体(intracellular receptor)两种类型。两种类型。配体闸门离子通道配体闸门离子通道(离子通道偶联受体)(离子通道偶联受体)G蛋白偶联受体蛋白偶联受体 生长因子类受体(酶偶联受体)生长因子类受体(酶偶联受体)细胞表面受体细胞表面受体 (cell surface receptor)细胞内受体细胞内受体(
10、intracellular receptor)胞浆受体胞浆受体核受体核受体v 受体的类型受体的类型v 离子通道偶联受体离子通道偶联受体(ion-channel-linked receptor)是一类自身为离子通道的受体,本身既有信号结合位是一类自身为离子通道的受体,本身既有信号结合位点,又是离子通道,其跨膜信号转导无需中间步骤。点,又是离子通道,其跨膜信号转导无需中间步骤。主要存在于神经、肌肉等可兴奋细胞,其信号分子为主要存在于神经、肌肉等可兴奋细胞,其信号分子为神经递质神经递质。离子通道型受体分为阳离子通道,如乙酰胆碱、谷氨离子通道型受体分为阳离子通道,如乙酰胆碱、谷氨酸和五羟色胺的受体,和
11、阴离子通道,如甘氨酸和酸和五羟色胺的受体,和阴离子通道,如甘氨酸和氨基丁酸的受体。氨基丁酸的受体。细胞表面受体细胞表面受体(cell surface receptor)神经递质通过与受体的结合而神经递质通过与受体的结合而改变通道蛋白的构象,导致离改变通道蛋白的构象,导致离子通道的开启或关闭,改变质子通道的开启或关闭,改变质膜的离子通透性,在瞬间将膜的离子通透性,在瞬间将胞胞外化学信号转换为电信号外化学信号转换为电信号,继,继而改变突触后细胞的兴奋性。而改变突触后细胞的兴奋性。离子通道型受体离子通道型受体乙酰胆碱受体结构模型乙酰胆碱受体结构模型 vv 离子通道受体介导胞外化学信号转变为电信号离子
12、通道受体介导胞外化学信号转变为电信号离子通道受体介导胞外化学信号转变为电信号离子通道受体介导胞外化学信号转变为电信号G蛋白偶联型受体(蛋白偶联型受体(G protein-linked receptor)G蛋白偶联受体指配体与细胞表面受体结合后激蛋白偶联受体指配体与细胞表面受体结合后激活偶联的活偶联的G蛋白,活性蛋白,活性G蛋白再激活产生第二信使蛋白再激活产生第二信使的酶类,通过产生第二信使引起细胞的生物学效应。的酶类,通过产生第二信使引起细胞的生物学效应。G蛋白偶联受体是一个连续蛋白偶联受体是一个连续7次跨膜的蛋白。次跨膜的蛋白。该受体该受体 氨基末端位于胞外,羧基末端位于胞内,跨膜部分为疏水
13、氨基末端位于胞外,羧基末端位于胞内,跨膜部分为疏水结构,由结构,由20-27个氨基酸残基组成。个氨基酸残基组成。受体受体胞外结构域胞外结构域识别胞外信号分子并与之结合,识别胞外信号分子并与之结合,胞内结构域胞内结构域与与G蛋蛋白耦联。通过与白耦联。通过与G蛋白耦联,调节相关酶活性,在细胞内产生第二信蛋白耦联,调节相关酶活性,在细胞内产生第二信使,从而将胞外信号跨膜传递到胞内。使,从而将胞外信号跨膜传递到胞内。G蛋白耦联型受体包括多种神经递质、肽类激素和趋化因子的受体,蛋白耦联型受体包括多种神经递质、肽类激素和趋化因子的受体,在味觉、视觉和嗅觉中接受外源理化因素的受体亦属在味觉、视觉和嗅觉中接受
14、外源理化因素的受体亦属G蛋白耦联型受蛋白耦联型受体。体。由由G蛋白耦联受体所介导的细胞信号通路主要包括:蛋白耦联受体所介导的细胞信号通路主要包括:cAMP信号通路和磷脂酰肌醇信号通路。信号通路和磷脂酰肌醇信号通路。G蛋白偶联受体的结构特征蛋白偶联受体的结构特征G蛋白耦联型受体为蛋白耦联型受体为7次跨膜蛋白次跨膜蛋白酶酶偶偶联联型型受受体体(enzyme linked receptor)分分为为两两类类,其其一一是是本本身身具具有有激激酶酶活活性性,如如肽肽类类生生长长因因子子(EGF,PDGF,CSF等等)受受体体;其其二二是是本本身身没没有有酶酶活活性性,但但可可以以连接非受体酪氨酸激酶,连
15、接非受体酪氨酸激酶,如细胞因子受体超家族。如细胞因子受体超家族。这这类类受受体体的的共共同同点点是是:通通常常为为单单次次跨跨膜膜蛋蛋白白;接接受配体后发生二聚化而激活,启动其下游信号转导。受配体后发生二聚化而激活,启动其下游信号转导。已已知知六六类类:受受体体酪酪氨氨酸酸激激酶酶、酪酪氨氨酸酸激激酶酶连连接接的的受受体体、受受体体酪酪氨氨酸酸磷磷脂脂酶酶、受受体体丝丝氨氨酸酸/苏苏氨氨酸酸激激酶酶受受体体鸟鸟苷苷酸酸环环化化酶酶、组组氨氨酸酸激激酶酶连连接接的的受受体体(与与细细菌的趋化性有关)。菌的趋化性有关)。酶偶联型受体酶偶联型受体酪氨酸激酶酪氨酸激酶酪酪氨氨酸酸激激酶酶可可分分为为三
16、三类类:受受体体酪酪氨氨酸酸激激酶酶,为为单单次次跨跨膜膜蛋蛋白白,在在脊脊椎椎动动物物中中已已发发现现50余余种种;胞胞质质酪酪氨氨酸酸激激酶酶,如如Src家家族族、Tec家家族族、ZAP70、家家族族、JAK家家族族等;等;核内酪氨酸激酶如核内酪氨酸激酶如Abl和和Wee。受受体体酪酪氨氨酸酸激激酶酶(receptor protein tyrosine kinases,RPTKs)是是单单次次跨跨膜膜蛋蛋白白,其其胞胞外外区区是是结结合合配配体体结结构构域域,配配体体是是可可溶溶性性或或膜膜结结合合的的多多肽肽或或蛋蛋白白类类激激素素,包包括括胰胰岛岛素素和和多多种种生生长长因因子子。胞胞
17、内内段段是是酪酪氨氨酸酸蛋蛋白白激激酶酶的的催催化化部部位位,并并具具有有自自磷磷酸酸化化位位点点。配配体体与与受受体体结结合合,导导致致受受体体二二聚化,二聚体内彼此相互磷酸化胞内段酪氨酸残基。聚化,二聚体内彼此相互磷酸化胞内段酪氨酸残基。v受体酪氨酸激酶受体酪氨酸激酶配配体体(如如EGF)在在胞胞外外与与受受体体结结合合并并引引起起构构象象变变化化,导导致致受受体体二二聚聚化化(dimerization)形形成成同同源源或或异异源源二二聚聚体体,在在二二聚聚体体内内彼彼此此相相互互磷磷酸酸化化胞胞内内段段酪酪氨氨酸酸残残基基,激激活活受受体体本本身身的的酪酪氨氨酸酸蛋蛋白白激激酶酶活活性性
18、。这这类类受受体体主主要有要有EGF、PDGF、FGF等,相应的配体一般为干扰素和生长因子等。等,相应的配体一般为干扰素和生长因子等。受体酪氨酸激酶的二聚化和自磷酸化受体酪氨酸激酶的二聚化和自磷酸化各类受体酪氨酸激酶(各类受体酪氨酸激酶(EGF:表皮生长因子;表皮生长因子;IGF:类胰岛素生长因类胰岛素生长因子;子;NGF:神经生长因子;神经生长因子;FGF:成纤维细胞生长因子;成纤维细胞生长因子;PDGF:血血小板生长因子;小板生长因子;VEGF:血管内皮生长因子;)血管内皮生长因子;)膜表面受体的三种类型膜表面受体的三种类型亲水性化学信号分子亲水性化学信号分子(包括神经递质、蛋白(包括神经
19、递质、蛋白激素、生长因子等)不激素、生长因子等)不能直接进入细胞,只能能直接进入细胞,只能通过通过膜表面的特异受体膜表面的特异受体传递信号,使靶细胞产传递信号,使靶细胞产生效应。生效应。第二节第二节 膜表面受体介导的信号转导膜表面受体介导的信号转导亲亲水水性性化化学学信信号号分分子子(包包括括神神经经递递质质、蛋蛋白白激激素素、生生长长因因子子等等)不不能能直直接接进进入入细细胞胞,只只能能通通过过膜膜表表面面的的特特异异受受体体传递信号,使靶细胞产生效应。传递信号,使靶细胞产生效应。膜表面受体主要有三类:膜表面受体主要有三类:离子通道型受体(离子通道型受体(ion-channel-linke
20、d receptor););G蛋白耦联型受体(蛋白耦联型受体(G-protein-linked receptor););酶耦联的受体(酶耦联的受体(enzyme-linked receptor)。)。第第一一类类存存在在于于可可兴兴奋奋细细胞胞。后后两两类类存存在在于于大大多多数数细细胞胞,在在信信号号转转导导的的早早期期表表现现为为激激酶酶级级联联(kinase cascade)事事件件,即即为为一一系系列列蛋蛋白白质质的的逐逐级级磷磷酸酸化化,籍籍此此使使信信号号逐逐级级传传送和放大。送和放大。v 由由G蛋白耦联型受体所介导的细胞信号通路:蛋白耦联型受体所介导的细胞信号通路:cAMP信号通
21、路和磷脂酰肌醇信号通路。信号通路和磷脂酰肌醇信号通路。v 由酶偶联受体所介导的细胞信号通路:由酶偶联受体所介导的细胞信号通路:酪氨酸激酶受体激活的酪氨酸激酶受体激活的Ras信号通路和细胞因子受体超信号通路和细胞因子受体超家族的家族的JAK-STAT信号通路信号通路。v 概念:概念:G蛋白(蛋白(G protein)全称为鸟苷酸结合蛋白全称为鸟苷酸结合蛋白(guanine nucleotide-binding protein),),通常指信号转通常指信号转导途径中与受体偶联的鸟苷酸结合蛋白。导途径中与受体偶联的鸟苷酸结合蛋白。v G蛋白特点:蛋白特点:位于质膜胞质侧,由位于质膜胞质侧,由、三个亚
22、基组成;三个亚基组成;具有结合具有结合GDP 和和GTP的能力,并具有的能力,并具有GTP酶活性;酶活性;G蛋白在信号转导过程中起着蛋白在信号转导过程中起着分子开关分子开关的作用,当的作用,当亚基与亚基与GDP结合时处于关闭状态,与结合时处于关闭状态,与GTP结合时处于开启状态,结合时处于开启状态,亚基具有亚基具有GTP酶活性,能催化所结合的酶活性,能催化所结合的ATP水解,恢复无活性的三聚体状态。水解,恢复无活性的三聚体状态。构象改变可以进一步激活效应蛋白,实现细胞外信号传递到细构象改变可以进一步激活效应蛋白,实现细胞外信号传递到细胞内的过程。胞内的过程。由由G蛋白耦联型受体所介导的细胞信号
23、通路蛋白耦联型受体所介导的细胞信号通路G蛋白分子开关蛋白分子开关 (静息和激活状态静息和激活状态)G蛋白在信号蛋白在信号转导过程中起转导过程中起着着分子开关分子开关的的作用,当作用,当亚基亚基与与GDP结合时结合时处于关闭状态,处于关闭状态,与与GTP结合时结合时处于开启状态。处于开启状态。vThe structure and activation of G proteinsG蛋白分类:蛋白分类:刺激性刺激性G蛋白蛋白(Gs)抑制性抑制性G蛋白(蛋白(Gi)刺激性受体(刺激性受体(Rs)抑制性受体(抑制性受体(Ri)1、cAMP信号转导途径信号转导途径 在在cAMP信号途径中,细胞外信号与相应
24、信号途径中,细胞外信号与相应受体结合,调节腺苷酸环化酶活性,通过受体结合,调节腺苷酸环化酶活性,通过第二信使第二信使cAMP水平的变化,将细胞外信水平的变化,将细胞外信号转变为细胞内信号。号转变为细胞内信号。刺激性刺激性G蛋白复蛋白复合体合体(Gs)抑制性抑制性G蛋白复合蛋白复合体体(Gi)刺激型激素刺激型激素受体受体 (Rs)抑制型激素抑制型激素受体受体 (Ri)腺苷酸环化酶腺苷酸环化酶 (AC)v cAMP信号通路的组成信号通路的组成cAMP信号途径的组成信号途径的组成 刺激型激素受体刺激型激素受体(Rs)或抑制型激素受体(或抑制型激素受体(Ri)刺激型调节蛋白刺激型调节蛋白(Gs)或抑制
25、型调节蛋白(或抑制型调节蛋白(Gi)腺苷酸环化酶腺苷酸环化酶(Adenylyl cyclase,AC)蛋白激酶蛋白激酶A(Protein Kinase A,PKA)环腺苷酸磷酸二酯酶(环腺苷酸磷酸二酯酶(cAMP phosphodiesterase)激素受体(激素受体(R)的一般结构模式图,的一般结构模式图,7次穿膜。次穿膜。膜外有信号分子结合位点,膜内有与膜外有信号分子结合位点,膜内有与G-蛋白相作用的位点蛋白相作用的位点信号分子结信号分子结合位点合位点与与G-蛋白相蛋白相作用的位点作用的位点G蛋白偶联受体结构蛋白偶联受体结构腺苷酸环化酶结构腺苷酸环化酶结构腺苷酸环化酶腺苷酸环化酶(Aden
26、ylyl cyclase,AC)是相对分子量为是相对分子量为150KD的糖蛋白,跨膜的糖蛋白,跨膜12次。在次。在Mg2+或或Mn2+的存的存在下,腺苷酸环化酶催化在下,腺苷酸环化酶催化ATP生成生成cAMP。蛋白激酶蛋白激酶A结构结构蛋白激酶蛋白激酶A(Protein Kinase A,PKA):由两个催化亚基和两个调节亚基组由两个催化亚基和两个调节亚基组成。成。cAMP与调节亚基结合,改变调节亚基构象,使调节亚基和催化亚基解与调节亚基结合,改变调节亚基构象,使调节亚基和催化亚基解离,释放出催化亚基。活化的蛋白激酶离,释放出催化亚基。活化的蛋白激酶A催化亚基可使细胞内某些蛋白的丝催化亚基可使
27、细胞内某些蛋白的丝氨酸或苏氨酸残基磷酸化,于是改变这些蛋白的活性,进一步影响到相关基氨酸或苏氨酸残基磷酸化,于是改变这些蛋白的活性,进一步影响到相关基因的表达。因的表达。当细胞内产生第二信当细胞内产生第二信使使cAMP之后之后,过多的过多的cAMP可以在环腺苷可以在环腺苷酸磷酸二酯酶的作用酸磷酸二酯酶的作用下被降解下被降解,生成生成5-AMP,该酶起终止信该酶起终止信号的作用。号的作用。环腺苷酸磷酸二酯酶(环腺苷酸磷酸二酯酶(cAMP phosphodiesterase)cAMP的降解的降解激素配体激素配体+G蛋白受体蛋白受体Rs结合结合G蛋白受体蛋白受体Rs构象改变构象改变暴露出与暴露出与G
28、蛋白蛋白(Gs)结合的位点结合的位点激素激素-受体复合物与受体复合物与Gs结结合合Gs的的亚基构象改变亚基构象改变与与GDP结合力减弱结合力减弱,释放释放GDP同时同时,Mg 2+存在下存在下,复合物复合物结合结合GTP而活化而活化配体配体G蛋白偶蛋白偶联受体联受体 cAMP信号信号通路的信号传导过程通路的信号传导过程三聚体三聚体Gs蛋白解离出蛋白解离出亚基和亚基和基复合物基复合物,暴露出暴露出亚基与腺苷酸亚基与腺苷酸环化酶的结合位点环化酶的结合位点结合结合GTP的的亚基与腺苷酸环化酶亚基与腺苷酸环化酶结合,活化该酶结合,活化该酶,同时同时GTP水解水解.被被激活的腺苷酸环化酶激活的腺苷酸环化
29、酶(AC)催化细胞催化细胞中的中的ATP生成生成cAMP,cAMP作为第作为第二信使可以激活一系列靶酶二信使可以激活一系列靶酶,在细在细胞内引发一系列生理效应胞内引发一系列生理效应.亚基恢复原来的构象并导致与亚基恢复原来的构象并导致与AC解离,终止解离,终止AC的活化作用。的活化作用。亚基亚基与与亚基重新结合,使细胞回复到亚基重新结合,使细胞回复到静止状态。静止状态。Gs的的亚基构象改变与亚基构象改变与GDP结合结合力减弱力减弱,释放释放GDP同时同时,Mg 2+存在存在下下,复合物结合复合物结合GTP而活化而活化 cAMP signaling pathway vG-proteinactiva
30、tionandinactivationcycleRs-Gs-AC-cAMP途径途径当细胞受到激素刺激时当细胞受到激素刺激时:激素配体激素配体G蛋白受体蛋白受体Rs+G蛋白受体蛋白受体(Rs)构象改变暴露构象改变暴露出与出与G蛋白蛋白(Gs)结合的位点结合的位点激素激素-受体复受体复合物与合物与Gs结合结合Gs的的亚基构象改变亚基构象改变,与与GDP结合力减弱结合力减弱,释放释放GDPMg2+存在下存在下,复合物复合物结合结合GTP而活化而活化三聚体三聚体Gs蛋白解离出蛋白解离出亚基和亚基和基复合物基复合物暴露出暴露出亚基与腺苷酸环亚基与腺苷酸环化酶的结合位点化酶的结合位点结合结合GTP的的亚基
31、与腺苷酸亚基与腺苷酸环化酶结合,活化该酶环化酶结合,活化该酶.ATPcAMPcAMPcAMP作为第二信使激活一系列靶酶作为第二信使激活一系列靶酶,在细胞内引发一系列生理效应在细胞内引发一系列生理效应cAMP信号与糖原降解信号与糖原降解:不同细胞对不同细胞对cAMP信号途径的反应速度不同,信号途径的反应速度不同,在肌肉细胞在肌肉细胞1秒钟之内可启动糖原降解为葡糖秒钟之内可启动糖原降解为葡糖1-磷酸,而抑制糖原的磷酸,而抑制糖原的合成。合成。cAMP激活蛋白激活蛋白激酶激酶A激活磷酸激活磷酸化酶激酶化酶激酶激活糖原激活糖原磷酸化酶磷酸化酶催化糖原降解为催化糖原降解为葡萄糖葡萄糖1-1-磷酸磷酸受体
32、受体-G蛋白蛋白-AC-cAMP-蛋白激酶蛋白激酶-磷酸化磷酸化酶级联放大系统酶级联放大系统 催化靶蛋白磷酸化催化靶蛋白磷酸化ACcAMP激活蛋白激酶激活蛋白激酶A(PKA)激活其它酶激活其它酶引起生命活动的变化引起生命活动的变化ATP不同细胞对不同细胞对cAMP信号途径的反应速度不同:信号途径的反应速度不同:v在肌肉细胞,在肌肉细胞,1秒钟内可以启动糖原降解为葡萄糖秒钟内可以启动糖原降解为葡萄糖1-磷酸,而抑制糖原合成。磷酸,而抑制糖原合成。v在在某某些些分分泌泌细细胞胞,需需要要几几个个小小时时,激激活活的的PKA 进进入入细细胞胞核核,将将CRE结结合合蛋蛋白白磷磷酸酸化化,这这类类蛋蛋
33、白白通通常常是是cAMP反反应应元元件件结结合合蛋蛋白白(cAMP responsive element-binding protein,CREB)等等基基因因表表达达的的调调节节因因子子,激激活活后后的的CREB可可结结合合相相关关基基因因的的CRE区区,调调节节相相关关基基因因的的表表达达,表表达达的的蛋蛋白白质质产产物物对对细细胞胞产产生生各各种种生生物物学学效效应应。CRE(cAMP responsive element)是是DNA上上的的调调节节区区域域,序序 列列 为为 TGACGTCA。(CRE:CAMP Response Element,CREB:CRE Binding pro
34、tein)配体配体 受体受体与与G蛋白结合,激活蛋白结合,激活ACAC催化催化ATP产生第二产生第二信使信使cAMPcAMP激活激活PKAPKA使底物蛋白使底物蛋白CREB磷酸化磷酸化激活后的激活后的CREB可结可结合相关基因的合相关基因的CRE区,区,调控靶基因的表达调控靶基因的表达CRE:CAMP Response ElementCREB:CRE Binding protein霍霍乱乱毒毒素素是是由由霍霍乱乱杆杆菌菌产产生生的的肽肽;霍霍乱乱的的症症状状是是严严重重腹腹泻,患者脱水死亡。泻,患者脱水死亡。霍霍霍霍乱乱乱乱毒毒毒毒素素素素由由两两种种肽肽链链组组成成,其其一一是是ADPADP
35、核核核核糖糖糖糖转转转转移移移移酶酶酶酶。它它可可穿穿过过细细胞胞表表面面进进入入细细胞胞质质,催催化化胞胞内内NAD+的的ADP核核糖糖基基共共价价结结合合到到Gs的的亚亚基基的的修修饰饰位位点点上上;这这种种亚亚基基不不可可逆逆地地修修饰饰,使使它它可可与与GTP结结合合。但但丧丧失失了了GTP酶酶活活性性,GTP不不能能水水解解为为GDP,因因此此活活化化亚亚基基始始终终结结合合在在环化酶上,同样使其长久活化。环化酶上,同样使其长久活化。细细胞胞质质中中cAMP增增加加了了100倍倍以以上上,导导致致膜膜蛋蛋白白让让大大量量水分进入肠腔,造成严重腹泻。水分进入肠腔,造成严重腹泻。以以上上
36、事事实实的的阐阐明明不不仅仅弄弄清清了了霍霍乱乱疾疾病病机机制制,也也为为G蛋蛋白作用模型提供了进一步证据。白作用模型提供了进一步证据。与Rs-Gs-AC-cAMP途径相关的疾病途径相关的疾病 细菌毒素催化细菌毒素催化NAD+转移转移ADP-核糖基团到核糖基团到Gs蛋白蛋白亚基亚基ADP 核糖转移酶核糖转移酶Gs蛋白蛋白亚基亚基丧失了丧失了GTP酶活性,酶活性,GTP不能不能水解为水解为GDP,活化活化亚基始终亚基始终结合在结合在AC上,使其长久活化,上,使其长久活化,胞质中胞质中cAMP增加。增加。霍霍霍霍乱乱乱乱毒毒毒毒素素素素由由两两种种肽肽链链组组成成,其其一一是是ADPADP核核核核
37、糖糖糖糖转转转转移移移移酶酶酶酶。它它可可穿穿过过细细胞胞表表面面进进入入细细胞胞质质,催催化化胞胞内内NAD+的的ADP核核糖糖基基共共价价结结合合到到Gs的的亚亚基基的的修修饰饰位位点点上上;这这种种亚亚基基不不可可逆逆地地修修饰饰,使使它它可可与与GTP结结合合。但但丧丧失失了了GTP酶酶活活性性,GTP不不能能水水解解为为GDP,因因此此活活化化亚亚基基始始终终结结合合在在环环化化酶酶上,同样使其长久活化。上,同样使其长久活化。细细胞胞质质中中cAMP增增加加了了100倍倍以以上上,导导致致膜膜蛋蛋白白让让大大量量水水分分进进入入肠肠腔腔,造造成严重胶泻。成严重胶泻。以以上上事事实实的
38、的阐阐明明不不仅仅弄弄清清了了霍霍乱乱疾疾病病机机制制,也也为为G蛋蛋白白作作用用模模型型提提供供了了进一步证据。进一步证据。在磷脂酰肌醇信号通路中在磷脂酰肌醇信号通路中,胞外信号分子与细胞表面胞外信号分子与细胞表面G蛋蛋白耦联型受体结合,激活质膜上的磷脂酶白耦联型受体结合,激活质膜上的磷脂酶C(PLC),),使质膜上使质膜上4,5-二磷酸磷脂酰肌醇(二磷酸磷脂酰肌醇(PIP2)水解成水解成1,4,5-三磷酸肌醇(三磷酸肌醇(IP3)和二酰基甘油(和二酰基甘油(DG)两个第二两个第二信使,胞外信号转换为胞内信号,这一信号系统又称为信使,胞外信号转换为胞内信号,这一信号系统又称为“双信使系统双信
39、使系统”(double messenger system)。)。PIP2多数分布在质膜的胞质侧多数分布在质膜的胞质侧,少量分布在内质网的胞少量分布在内质网的胞质侧质侧,主要由内质网合成主要由内质网合成.2、磷脂酰肌醇信号转导途径、磷脂酰肌醇信号转导途径IP3、DG和和Ca2+信使体系的组成信使体系的组成受体受体磷脂磷脂酶酶(PLC)G蛋白蛋白磷脂酰肌醇信号通路磷脂酰肌醇信号通路 IP3、DG和和Ca 2+信使体系信使体系G蛋白偶蛋白偶联受体联受体配体配体+受体构象变化受体构象变化受体与受体与G蛋白结合蛋白结合G蛋白构象变化蛋白构象变化激活激活PLCPIP2DG+IP3(第二信使第二信使)Dou
40、ble Messenger systemIP3与内质网上的与内质网上的IP3配体门钙通道结合,开启钙通道,配体门钙通道结合,开启钙通道,使胞内使胞内Ca 2+浓度升高。激活各类依赖钙离子的蛋白。用浓度升高。激活各类依赖钙离子的蛋白。用Ca 2+载体离子霉素(载体离子霉素(ionomycin)处理细胞会产生类似处理细胞会产生类似的结果。的结果。DG结合于质膜上,可活化与质膜结合的蛋白激酶结合于质膜上,可活化与质膜结合的蛋白激酶C(Protein Kinase C,PKC)。)。PKC以非活性形式分布以非活性形式分布于细胞溶质中,当细胞接受刺激,产生于细胞溶质中,当细胞接受刺激,产生IP3,使使C
41、a 2+浓度浓度升高,升高,PKC便转位到质膜内表面,被便转位到质膜内表面,被DG活化,活化的活化,活化的PKC可以使蛋白质的丝氨酸可以使蛋白质的丝氨酸/苏氨酸残基磷酸化,使不同苏氨酸残基磷酸化,使不同的细胞产生不同的反应,如细胞分泌、肌肉收缩、细胞增的细胞产生不同的反应,如细胞分泌、肌肉收缩、细胞增殖和分化等。殖和分化等。DG的作用可用佛波醇酯(的作用可用佛波醇酯(phobol ester)模模拟。拟。Two intracellular pathways by which activated C-kinase can activate the transcription of specifi
42、c genes.In one(red arrows)C-kinase activates aphosphorylation cascade thatleadstothephosphorylationofapivotalproteinkinasecalledMAP-kinase,which in turnphosphorylates and activatesthe gene regulatory proteinElk-1.Elk-1isboundtoashortDNA sequence in associationwithanotherDNA-bindingprotein.In the oth
43、er pathway(green arrows)C-kinaseactivationleadstothephosphorylationofIk-B,whichreleasesthegeneregulatory protein NF-kB sothat it can migrate into thenucleusandactivatethetranscriptionofspecificgenes.MAPK为有丝分裂原活化蛋白激酶(为有丝分裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK),),属丝氨酸属丝氨酸/苏氨酸残基酶苏氨酸残基酶v IP3-Ca2
44、+pathway and DG-PKC pathwayElevation of cytosolic Ca2+via the IP signaling pathwaySignalsGPLR GP PLC IP3 and DAG(twin signals).IP3 IP3 receptor(Ca2+channel,located at the surface of sER)Elevation of cytosolic Ca2+;DAG activates PKC to phosphoralate Ser and Thr on target proteins.Calcium binds to cal
45、cium-binding proteins(CaM)which affects other proteins.AlfredG.Gilman MartinRodbellTheNobelPrizeinPhysiologyorMedicine1994fortheirdiscoveryofG-proteinsandtheroleoftheseproteinsinsignaltransductionincells 3、酶偶联受体介导的受体信号通路酶偶联受体介导的受体信号通路 酶偶联受体分为两种类型:酶偶联受体分为两种类型:本身具有激酶活性本身具有激酶活性,如,如EGF,PDGF,CSF等生长因子类等生长
46、因子类的受体本身具有的受体本身具有酪氨酸激酶活性,称为受体酪氨酸激酶酪氨酸激酶活性,称为受体酪氨酸激酶(receptor protein tyrosine kinases,RPTKs),),是单是单次跨膜蛋白,配体(如次跨膜蛋白,配体(如EGF等多种生长因子)与受体结等多种生长因子)与受体结合合,导致受体二聚化,二聚体内彼此相互磷酸化胞内段导致受体二聚化,二聚体内彼此相互磷酸化胞内段酪氨酸残基。酪氨酸残基。本身没有酶活性本身没有酶活性,但可以连接非受体酪氨酸激酶,完,但可以连接非受体酪氨酸激酶,完成信号从胞外到胞内的传递,如细胞因子受体超家族。成信号从胞外到胞内的传递,如细胞因子受体超家族。这
47、这类类受受体体的的共共同同点点是是:通通常常为为单单次次跨跨膜膜蛋蛋白白;接接受配体后发生二聚化而激活,启动其下游信号转导。受配体后发生二聚化而激活,启动其下游信号转导。已知六类:已知六类:受体酪氨酸激酶受体酪氨酸激酶、酪氨酸激酶连接的受体、酪氨酸激酶连接的受体、受体酪氨酸磷脂酶、受体酪氨酸磷脂酶、受体丝氨酸受体丝氨酸/苏氨酸激酶苏氨酸激酶受体鸟苷酸环化酶、受体鸟苷酸环化酶、组氨酸激酶连接的受体(与细菌的趋化性有关)。组氨酸激酶连接的受体(与细菌的趋化性有关)。酶偶联型受体(酶偶联型受体(enzyme linked receptor)酪氨酸激酶酪氨酸激酶酪酪氨氨酸酸激激酶酶可可分分为为三三类类
48、:受受体体酪酪氨氨酸酸激激酶酶,为为单单次次跨跨膜膜蛋蛋白白,在在脊脊椎椎动动物物中中已已发发现现50余余种种;胞胞质质酪酪氨氨酸酸激激酶酶,如如Src家家族族、Tec家家族族、ZAP70、家家族族、JAK家家族族等;等;核内酪氨酸激酶如核内酪氨酸激酶如Abl和和Wee。受受体体酪酪氨氨酸酸激激酶酶(receptor protein tyrosine kinases,RPTKs)是是单单次次跨跨膜膜蛋蛋白白,其其胞胞外外区区是是结结合合配配体体结结构构域域,配配体体是是可可溶溶性性或或膜膜结结合合的的多多肽肽或或蛋蛋白白类类激激素素,包包括括胰胰岛岛素素和和多多种种生生长长因因子子。胞胞内内段
49、段是是酪酪氨氨酸酸蛋蛋白白激激酶酶的的催催化化部部位位,并并具具有有自自磷磷酸酸化化位位点点。配配体体与与受受体体结结合合,导导致致受受体体二二聚化,二聚体内彼此相互磷酸化胞内段酪氨酸残基。聚化,二聚体内彼此相互磷酸化胞内段酪氨酸残基。v 受体酪氨酸激酶受体酪氨酸激酶v 受体酪氨酸激酶的结构与组成受体酪氨酸激酶的结构与组成受体:生长因子受体:生长因子结合部位,胞外结合部位,胞外跨膜跨膜螺旋:螺旋:与膜结合部位与膜结合部位胞质结构域:酪氨酸胞质结构域:酪氨酸激酶酪氨酸激酶酪氨酸“尾尾”胞外区是结合配体的部位,胞外区是结合配体的部位,配体是可溶性或膜结合的多肽或蛋白配体是可溶性或膜结合的多肽或蛋白
50、类激素,包括胰岛素和多种生长因子。类激素,包括胰岛素和多种生长因子。胞内段是酪氨酸蛋白激酶的催化部位,具有酪氨酸激酶的活性胞内段是酪氨酸蛋白激酶的催化部位,具有酪氨酸激酶的活性。Six subfamilies of receptor tyrosine kinases.1.Epidermal growth factor(EGF)receptor,2.insulin and insulin-like GF(IGF-1)receptor,3.Nerve growth factor(NGF)receptor,4.Platelet-derived growth factor(PDGF)receptor,
51、5.Fibroblast growth factor(FGF)receptor,6.vascular endothelial factor(VEGF)receptor.几种生长因子受体几种生长因子受体均属于受体酪氨酸激酶均属于受体酪氨酸激酶v 信号转导过程信号转导过程激素与受体结合激素与受体结合受体构象变化,相邻受体构象变化,相邻受体靠近受体靠近胞内酪氨酸互相磷酸胞内酪氨酸互相磷酸化化自磷酸化自磷酸化激活酪氨酸蛋白激酶激活酪氨酸蛋白激酶激活含有激活含有SH2结构域的结构域的蛋白蛋白受体酪氨酸激酶的二聚化和自磷酸化受体酪氨酸激酶的二聚化和自磷酸化v 酪氨酸激酶受体激活的酪氨酸激酶受体激活的Ras
52、 信号通路信号通路配体与受体结合配体与受体结合 激活激活酪氨酸蛋白激酶酪氨酸蛋白激酶 接头蛋白接头蛋白Grb2通过通过SH2与受体的磷酸酪氨酸与受体的磷酸酪氨酸残基结合,再通过残基结合,再通过SH3与与鸟苷酸交换因子鸟苷酸交换因子 Sos结合,结合,Sos与膜上的与膜上的Ras接触,从而接触,从而活化活化Ras。Ras蛋蛋白白与与Raf的的N端端结结构构域域结结合合并并使使其其激激活活,Raf是是丝丝氨氨酸酸/苏苏氨氨酸酸(Ser/Thr)蛋蛋白白激激酶酶(又又称称MAPKKK)。活活化化的的Raf结结合合并并磷磷酸酸化化另另一一种种蛋蛋白白激激酶酶MAPKK,使使其其活活化化。MAPKK又又
53、使使MAPK的的苏苏氨氨酸酸和和酪酪氨氨酸酸残残基基使之激活。使之激活。MAPK为为 有有 丝丝 分分 裂裂 原原 活活 化化 蛋蛋 白白 激激 酶酶(mitogen-activated protein kinase,MAPK),属属丝丝氨氨酸酸/苏苏氨氨酸酸残残激激酶酶。活活化化的的MAPK进进入入细细胞胞核核,可可使使许许多多转转录录因因子子活活化化,如如将将Elk-1激激活活,促促进进c-fos,c-jun的的表表达。达。v 酪氨酸激酶受体激活的酪氨酸激酶受体激活的Ras信号通路信号通路受体酪氨酸激酶(受体酪氨酸激酶(RPTK)结合结合信号分子,形成二聚体,并发生信号分子,形成二聚体,并
54、发生自磷酸化而活化,活化的自磷酸化而活化,活化的RPTK激活激活RAS,由活化的由活化的RAS引起蛋引起蛋白激酶的磷酸化级联反应白激酶的磷酸化级联反应 RPTK-Ras信信号号通通路路可可概概括括如如下:下:配配体体RPTK(受受体体酪酪氨氨酸酸激激酶酶)adaptorGEF(鸟鸟苷苷酸酸交交换换因因子子如如:Sos)RasRaf(MAPKKK)MAPKKMAPK进进入入细细胞胞核核转录因子转录因子基因表达。基因表达。Ras蛋白要释放蛋白要释放GDP,才能与才能与GTP结合后激活,结合后激活,GDP的释放需要的释放需要鸟苷酸交鸟苷酸交换因子(换因子(GEF,如如Sos)参与;参与;Ras本身的
55、本身的GTP酶活性不强,需要酶活性不强,需要GTP酶活酶活化蛋白(化蛋白(GAP)的参与,使的参与,使Ras结合的结合的GTP水解而失活,水解而失活,GAP具有具有SH2结构结构域可直接与活化的受体结合。域可直接与活化的受体结合。v MAP-kinase serine/threonine phosphorylation Pathway activated by Ras激激活活的的Ras蛋蛋白白与与MAPKKK的的N端端结结构构域域结结合合并并使使其其激激活活,MAPKKK 是是 丝丝 氨氨 酸酸/苏苏 氨氨 酸酸(Ser/Thr)蛋白激酶(又称蛋白激酶(又称Raf)激激活活的的MAPKKK与与
56、另另一一种种蛋蛋白白激酶激酶MAPKK结合并使其活化。结合并使其活化。活活化化的的MAPKK又又使使MAPK的的苏苏氨氨酸酸和和酪酪氨氨酸酸残残基基磷磷酸酸化化使使之之激激活。活。MAPK为为有有丝丝分分裂裂原原活活化化蛋蛋白白激激酶酶(mitogen-activated protein kinase,MAPK),属属丝丝氨氨酸酸/苏苏氨氨酸酸残残基基酶酶。活活化化的的MAPK进进入入细细胞胞核核,可可使使许许多多转转录录因因子子活活化化,进进而而影影响响某某些些基基因因的的表表达达,还还可可以以改改变某些蛋白的活性。变某些蛋白的活性。vRPTK-Ras signaling pathwayv
57、此此类类受受体体属属于于酪酪氨氨酸酸激激酶酶连连接接的的受受体体(tyrosine kinase associated receptor)。细细胞胞因因子子(cytokine),如如:白白介介素素(IL)、干干扰扰素素(IFN)、集集落落刺刺激激因因子子(CSF)、生生长长激激素素(GH)、转转移移生生长长因因子子(TGF)等,在造血细胞和免疫细胞通讯上起作用。等,在造血细胞和免疫细胞通讯上起作用。v细细胞胞因因子子受受体体为为单单次次跨跨膜膜蛋蛋白白,均均由由两两个个或或多多个个多多肽肽链链组组成成,本本身身不不具具有有酶酶活活性性,但但与与配配体体结结合合后后发发生生二二聚聚化化而而激激活
58、活,与与胞胞内内酪酪氨氨酸酸蛋蛋白白激激酶酶(如如,JAK)连连接接,其信号途径为其信号途径为JAKSTAT。细胞因子受体超家族的细胞因子受体超家族的JAK-STAT信号通路信号通路v JAK(just another kinase或或janus kinase)是一类是一类非受体酪氨酸激酶家族非受体酪氨酸激酶家族,属于,属于胞质胞质酪氨酸激酶,酪氨酸激酶,已发已发现四个成员,即现四个成员,即JAK1、JAK2、JAK3 和和TYK1,每一每一个成员都与一个特异的细胞因子受体相连接个成员都与一个特异的细胞因子受体相连接。其结构。其结构不含不含SH2、SH3,C段具有两个相连的激酶区。段具有两个相
59、连的激酶区。v JAK的底物为的底物为STAT,即信号转导子和转录激活子即信号转导子和转录激活子(signal transducer and activator of transcription,STAT),),具有具有SH2和和SH3两类结构域。两类结构域。STAT被被JAK磷酸化后发生二聚化,然后穿过核膜进入核内调节相磷酸化后发生二聚化,然后穿过核膜进入核内调节相关基因的表达,这条信号通路称为关基因的表达,这条信号通路称为JAK-STAT途径途径。v 体内约超过体内约超过30种细胞因子和激素通过与相应的细胞种细胞因子和激素通过与相应的细胞因子受体结合激活因子受体结合激活Jak-STAT信号
60、通路。信号通路。TheJak-STATsignalingpathwayavtivatedby-interferon.Providingafasttracktothenucleus.MBC885:15-63JAK-STAT信信号号途途径径可可概概括括如下:如下:配配体体(Cytokine)与与受受体体结合导致受体二聚化;结合导致受体二聚化;二聚化受体激活二聚化受体激活JAK;JAK将将STAT磷酸化;磷酸化;STAT形形成成二二聚聚体体,暴暴露露出入核信号;出入核信号;STAT进进入入核核内内,调调节节基基因表达。因表达。第三节第三节 胞内受体介导的信号传导胞内受体介导的信号传导 细细胞胞内内受
61、受体体的的本本质质是是激激素素激激活活的的基基因因调调控控蛋蛋白白。在在细细胞胞内内,受受体体与与抑抑制制性性蛋蛋白白(如如Hsp90)结结合合形形成成复复合合物物,处处于于非非活活化化状状态态。配配体体(如如皮皮质质醇醇)与与受受体体结结合合,将将导导致致抑抑制制性性蛋蛋白白从从复复合合物物上上解解离离下来,从而使受体暴露出下来,从而使受体暴露出DNA结合位点而被激活。结合位点而被激活。这这类类受受体体一一般般都都有有三三个个结结构构域域:位位于于C端端的的激激素素结结合合位位点点,位位于于中中部部富富含含Cys、具具有有锌锌指指结结构构的的DNA或或Hsp90结结合合位位点点,以以及及位位
62、于于N端端的的转转录录激激活结构域。活结构域。胞内受体结构胞内受体结构甾甾类类激激素素分分子子是是化化学学结结构构相相似似的的亲亲脂脂性性小小分分子子,分分子子相相对对质质量量为为300Da左左右右,可可以以通通过过简简单单扩扩散散跨跨越越质质膜膜进入细胞内。进入细胞内。每每种种类类型型的的甾甾类类激激素素与与细细胞胞质质内内各各自自的的受受体体蛋蛋白白结结合合,形形成成激激素素-受受体体复复合合物物,并并能能穿穿过过核核孔孔进进入入细细胞胞核核内内,激激素素和和受受体体的的结结合合导导致致受受体体蛋蛋白白构构象象的的改改变变,提提高高了了受受体体与与DNA的的结结合合能能力力,激激活活的的受
63、受体体通通过过结结合合于于特特异异的的DNA序列调节基因表达序列调节基因表达。受受体体与与DNA序序列列的的结结合合已已得得到到实实验验证证实实,结结合合序序列列是是受受体体依依赖赖的的转转录录增增强强子子,这这种种结结合合可可增增加加某某些些相相邻邻基基因的转录水平。因的转录水平。v 一氧化氮(一氧化氮(NO)信号信号NO是另一种可进入细胞内部的信号分子,能快速透过是另一种可进入细胞内部的信号分子,能快速透过细胞膜,作用于邻近细胞。细胞膜,作用于邻近细胞。RFurchgott等三位美国等三位美国科学家因发现科学家因发现NO作为信号分子而获得作为信号分子而获得1998年诺贝尔医年诺贝尔医学与生
64、理学奖。学与生理学奖。血血管管内内皮皮细细胞胞和和神神经经细细胞胞是是NO的的生生成成细细胞胞,NO的的生生成成由由一一氧氧化化氮氮合合酶酶(nitric oxide synthase,NOS)催催化化,以以L-精精氨氨酸酸为为底底物物,生生成成NO和和L-瓜瓜氨氨酸酸。NO没没有有专专门门的的储储存存及及释释放放调调节节机机制制,靶靶细细胞胞上上NO的的多多少少直直接与接与NO的合成有关。的合成有关。血血管管内内皮皮细细胞胞接接受受乙乙酰酰胆胆碱碱,引引起起胞胞内内Ca2+浓浓度度升升高高,激激活活一一氧氧化化氮氮合合酶酶,细细胞胞释释放放NO,NO扩扩散散进进入入平平滑滑肌肌细细胞胞,与与
65、胞胞质质鸟鸟苷苷酸酸环环化化酶酶(GTP-cyclase,GC)活活性性中中心心的的Fe2结结合合,改改变变酶酶的的构构象象,导导致致酶酶活活性性的的增强和增强和cGMP合成增多。合成增多。cGMP可可降降低低血血管管平平滑滑肌肌中中的的Ca2+离离子子浓浓度度。引引起起血血管管平平滑滑肌肌的的舒舒张张,血血管管扩扩张张、血血流流通畅。通畅。硝硝酸酸甘甘油油治治疗疗心心绞绞痛痛具具有有百百年年的的历历史史,其其作作用用机机理理是是在在体体内内转转化化为为NO,可可舒舒张张血血管管,减减轻心脏负荷和心肌的需氧量轻心脏负荷和心肌的需氧量。NO的作用机制的作用机制鸟苷酸环化酶鸟苷酸环化酶NO的作用机
66、制的作用机制乙酰胆碱乙酰胆碱血管内皮血管内皮胞内胞内Ca2+浓度升高浓度升高一氧化氮合酶一氧化氮合酶NO平滑肌细胞平滑肌细胞与与胞质胞质鸟苷酸环化酶结合鸟苷酸环化酶结合cGMP合成增多合成增多血管平滑肌细胞的血管平滑肌细胞的Ca2+离子浓度下降离子浓度下降平滑肌舒平滑肌舒张张血管扩张、血流通畅。血管扩张、血流通畅。细胞膜的细胞膜的信息传递信息传递功能功能基本概念基本概念G蛋白偶联的蛋白偶联的信号转导途径信号转导途径配体配体受体受体离子通道偶联受体离子通道偶联受体G蛋白偶联受体蛋白偶联受体酶偶联受体酶偶联受体CAMP信号转导途径信号转导途径磷脂酰肌醇信号转导途径磷脂酰肌醇信号转导途径脂溶性配体脂溶性配体 膜受体膜受体总结总结水溶性配体水溶性配体 胞内受体胞内受体 G蛋白蛋白 G蛋白偶联受体蛋白偶联受体 腺苷酸环化酶腺苷酸环化酶 蛋白激酶蛋白激酶A第二信使第二信使CAMP G蛋白蛋白 G蛋白偶联受体蛋白偶联受体 磷脂酶磷脂酶C 蛋白激酶蛋白激酶C第二信使第二信使DG+IP3酶偶联受体介导的酶偶联受体介导的信号转导途径信号转导途径酪氨酸激酶受体激活的酪氨酸激酶受体激活的Ras信号通路信号通路
- 温馨提示:
1: 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
2: 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
3.本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
5. 装配图网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
