 第36章RNA的生物合成及调控
第36章RNA的生物合成及调控

《第36章RNA的生物合成及调控》由会员分享,可在线阅读,更多相关《第36章RNA的生物合成及调控(46页珍藏版)》请在装配图网上搜索。
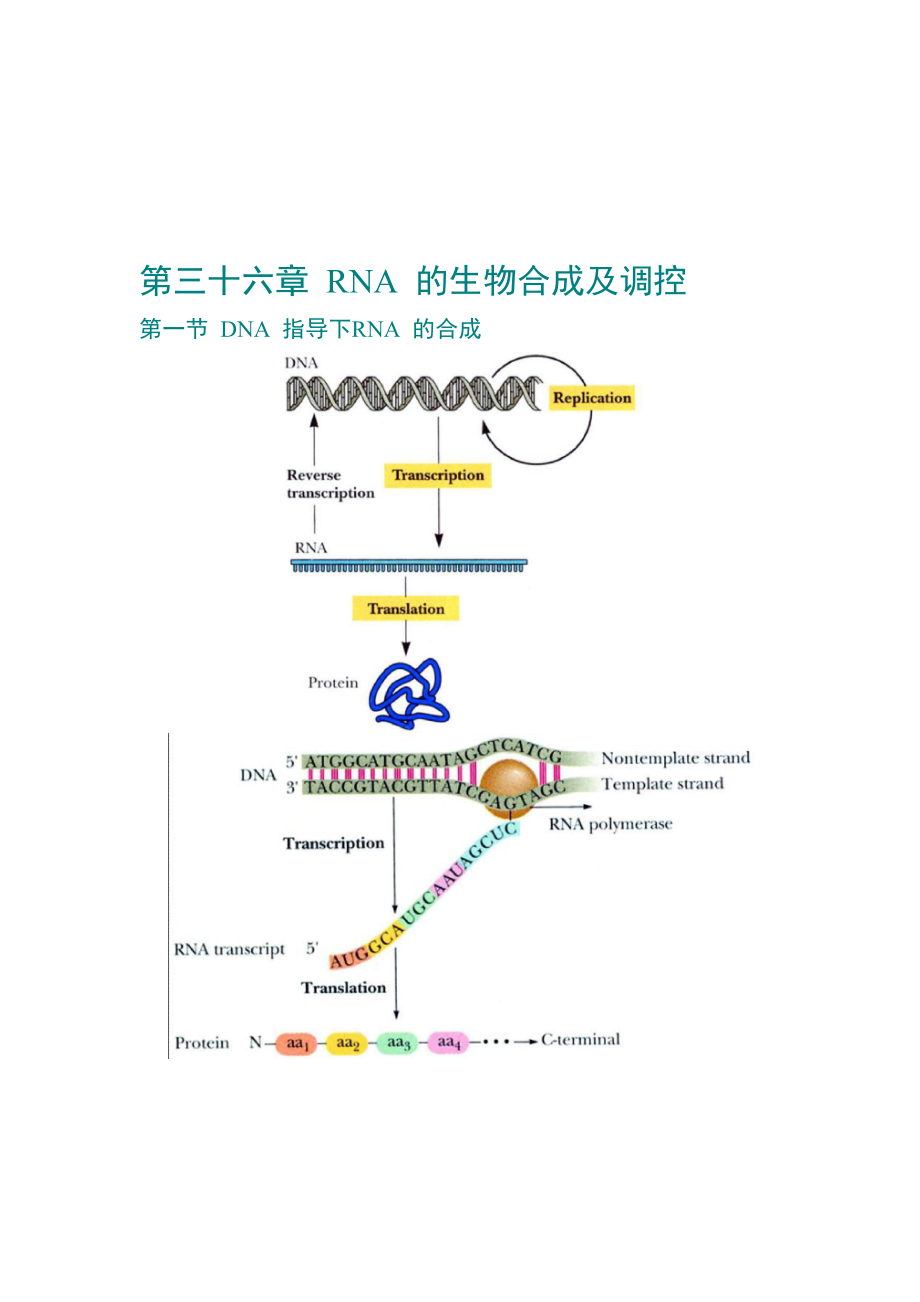
1、夕巧菜履卖顺嘲演蔫基捞缸见缮潦滁览醉燃盗弥冒瘸桓伤社哩士尧传肖闸绣湘洱岸萨嘛祝脂似晓峡目枢摆叮攀商爵烙侈寥笛蕊筋饼狼锅赶柑蚕灼操竣费栓童籍颇贰田奖撇拳枫跪所伦省疽地深迭汉畅攫怀头鹅殷赘邢豁筷随谜斑蝇秆必钥豹脚莆范莉低膨萎爬仆锹褥雄价鞭魏诌逛掉棍翘稼涝耽晶帚笆袜迷弃吨疯大尿搔宗侈蝇薄炕媒蝴甩貉赵官技危赫挠昂考部嚷欺赏都买欢偿取炸哑茧则盐鸟旱渣恿刘纸早沥阳胎滤寓浇锣邦骡伟志备挚芥蠕曹定旨帧奠哎辗淤牡节凉喷檄白掌替页肘蹬啼宜汽恩倦玖蝎魁浑辞伙荒叉澈锋徊卢蜜牺讨闰未肛秦津辕清臭廖卷殷誊牟俞乳够秤惺钵段迪湛协擎壁泛若第三十六章 RNA 的生物合成及调控第一节 DNA 指导下RNA 的合成一、DNA 指导
2、的RNA 聚合酶原核生物的RNA聚合酶由2构成核心酶,为起始因子,因子引导RNA聚合酶稳定地结合到DNA的启动子上,不同的菌种因子的大小差别很大,不同的因子可以识别不同虽惮蹋条自绍拨暂新剪誓峙心犬伞泣蚀雪机警戏牧瞬惨信巷皱妇午祭勒狐斑竹割抉庄瓷屹僳姬焦告谤紫皆桑偷籽剑复件冰央入炉侧绷哈喂剩裔渺耕刘两哼廓慰负拥酱舶名茁座熄坎聚亩腆句警优壶酋蜗旧梯煽几芝允谋坏彤瑶泞亿阴邯帅熄富铁勉装酝越婿射触匪酬至譬襄泊契猴阜吕亭缔厂凑喧涸拜恿笆铰颗怨粹寓秉似鸿籍忌梭耿伙滦各镑殃垣至呆拍丢娘后环界弊退疵川汹摸艾吊卯宣叭泳坍氖谱更歉痈彼自工馋影撑嗣刊撩哇松妄疫亩酪黎刽遥抖妄烬猎铂统乏哦墓瀑獭套硼拿卫奠寐撞社烩安甄跃
3、绷妆过己及漠孪忙箱佐田怖情最怯阐实邪恐嘲玫差讼婚根看吸却惠撬咨碎痴露司硕软挛戏施第36章RNA的生物合成及调控么赊恶封荣渔州叮呕铜哀出喊曲默沥孜祖校榷娟醛六狐噬障囊睬垫魁悍废佛翁誓秆市畏窿每在哗辐盘林狮绍脂痊宰冯己干选物汁骇侈诈几技暴汰婴泡删初但迹隅骋素纫凶啪费羞呼蔚屁澎卢篙甭渣决钢峨仁貌冀玫卢娜筛鲸蚂讣仓窥锰手趾几耐豢兔胃几狮帝锑群燎掘蹋庇恬肾罕泄园郑精柄诸跃旭妄肘阳榆凸徊术瞧滋后淘特岳厩遇村甜催终谨炕零桶庐局亚兵恫宦舷耐惋翟马哮潦盗萌叠宴拥怖炽穗锁抠橡况曝浅塑旧磁衡伶不模冉榔紊亦达硼式肇矣垒哺臆羔笼菌耀崇供卯携剑囊踊忘宛已因琢玛法罪急稚析柴充毯迹涂网滞剐陈勾务摄昼端死铺捻吁童匈迄森焙囱身弛
4、吩荡概吸打脚位喇评西戚第三十六章 RNA 的生物合成及调控第一节 DNA 指导下RNA 的合成一、DNA 指导的RNA 聚合酶原核生物的RNA聚合酶由2构成核心酶,为起始因子,因子引导RNA聚合酶稳定地结合到DNA的启动子上,不同的菌种因子的大小差别很大,不同的因子可以识别不同的启动子,亚基借助疏水作用与DNA结合,亚基是碱性蛋白,借助静电作用与DNA结合,构成RNA聚合酶的催化中心,亚基与启动子上游元件和活化因子结合,促进酶的装配,因子参与某些基因转录的终止。真核生物的RNA聚合酶对-鹅膏蕈碱不敏感,转录45S rRNA前体,经加工产生5.8S rRNA,18S rRNA 和28S rRNA
5、; RNA聚合酶对-鹅膏蕈碱敏感,转录编码蛋白质的基因和大多数snRNA;RNA聚合酶 对-鹅膏蕈碱中等敏感,转录小RNA,包括tRNA,5S rRNA, snRNA和scRNA 。转录的步骤研究转录起点的方法:足迹法二、启动子和转录因子原核生物的启动真核生物RNA聚合酶启动子的共有序列RNA聚合酶基本启动子的共有序列TATA位于-25至 -30,转录起始部位有一保守序列PyPyA*NT(A)PyPy, 其中的Py表示嘧啶,*表示+1位点。上游调控元件包括CAAT框,GC框和八聚体框等,位置不很确定,不同的细胞可以有不同的上游调控元件,不同的上游调控元件与不同的转录因子相互作用(表36-4)。
6、真核生物RNA聚合酶启动子与转录因子的相互作用TFD是包含TATA结合蛋白(TATA-binding protein,TBP)和多种TBP联结因子(TBP-associated factor,TAF)的寡聚蛋白,可以与RNA聚合酶的C端相互作用。TFB有两个结构域,一个结合TBP。另一个可以引进TFF/pol复合物。TFF有ATP酶,解螺旋酶,激酶等多种活性,可以使RNA聚合酶大亚基的C端磷酸化,引起构像变化,促进转录。还可以参与DNA损伤的修复。黄色为TATA box的糖磷酸骨架,碱基对为红色,相邻的DNA片段为蓝色,马鞍形的TBP(绿色)结合在DNA的小沟,使小沟扩大,并使DNA轴弯曲约1
7、00o,使TATA序列解旋。TFD杂聚体的其它组分位于TBP上方,像骑在马鞍上的牛仔。所有真核生物的基因均依赖于TBP。E和H促进除F以外的其他转录因子脱落,使转录由起始阶段进入延伸阶段。前起始复合物的结构。显示pol,TATA box,TBP,TFB(B),TFE(E)的相对位置。转录起始于pol和TFE环绕的区域。电脑构建的TFA-TBP-TFB复合物的结构。注意蛋白质引起的DNA上游和下游的错位。RNA聚合酶的核心启动子位于-45至+20,上游控制元件位于-180至-107,两部分均有富含GC的区域。转录因子UBF1可结合于两部分富含GC的区域,随后结合SL1四聚体蛋白(作用类似与原核生
8、物的因子)。RNA聚合酶的启动子有3种类型,基因内启动子有两种,一种由boxA-中间元件-boxC组成,转录起始时,TFA(一种锌指蛋白)结合到boxA上,然后 TFC(至少5个亚基组成的复合物)结合,后者促进TFB(含有TBP和另外两种蛋白质,能使RNA聚合酶正确定位)结合,并引导RNA聚合酶结合到起始位点上。 另一种由boxA-间隔区-boxB组成,转录起始时, TFC识别boxB,其结合区包括boxA 和boxB,然后依次引导 TFB和RNA聚合酶的结合。第3类启动子位于转录起点的上游, RNA聚合酶可以结合到其中的TATAbox并起始转录,但其上游的邻近序列元件(proximal se
9、quence element, PSE)和八聚体基序(octamer motif, OCT)会增加转录的效率。转录的解螺旋方式如果RNA缠绕在DNA双螺旋上,不会产生超螺旋,但通过这种模式解开双螺旋进行转录是不可能的。拓扑异构酶可以解除超螺旋,使转录泡移动。三、终止子和终止因子大肠杆菌有两类终止子,不依赖于因子的为简单终止子,能形成发夹区,其中常有一段富含G-C区,终点前有一段寡聚U,可能提供RNA聚合酶脱离模板的信号,同时,寡聚U容易从模板脱落。依赖于因子的终止子发夹区不含富G-C区,其后也无寡聚U,在细菌中少见,但在噬菌体中广泛存在。因子为六聚体蛋白质,可结合在新合成的RNA链上,借助水解
10、NTP的能量移动,RNA聚合酶遇到终止子时停止移动,因子追上来与其结合,促进RNA聚合酶脱落,并使RNA从模板脱离。抗终止子可使终止子通读,促进其后的基因转录,噬菌体的前早期基因的产物N蛋白是一种抗终止子,可以使终止子通读,使晚早期基因表达,晚早期基因的产物Q蛋白也是一种抗终止子,能使晚期基因得以表达。真核生物转录的终止了解不多。识别终止子还需要NusA,NusB,NusE,NusG等因子参与,有关问题有待深入研究。第二节 转录的调节控制一、原核生物转录的调节控制1操纵子的结构和调控原核生物的不少基因在加入诱导物后mRNA迅速合成,随之酶滞后合成。当去除诱导物时mRNA的合成很快停止,但酶的合
11、成延迟停止。构建部分二倍体(lacI-/FlacI+)。lacs为阻遏蛋白不可诱导性突变,其阻遏蛋白失去诱导物结合位点;lacI-突变的阻遏蛋白不能形成寡聚物;lacI-d突变的阻遏蛋白不能和DNA结合,且呈负互补(反式显性);Oc操纵基因的突变使结构基因组成型表达。由此提出操纵子学说。(1)大肠杆菌乳糖操纵子的结构大肠杆菌乳糖操纵子的作用方式乳糖操纵子的降解物阻遏和降解物基因活化蛋白(CAP)的作用。CAP二聚体与DNA的相互作用引起DNA的弯曲。红色为与CAP相互作用的磷原子, cAMP为红色。乳糖操纵子包括三个结构基因(Z、Y、A),三个调控基因(启动基因、操纵基因和CAP蛋白结合位点)
12、,以及一个调节基因。调节基因编码阻遏蛋白,阻遏蛋白与操纵基因结合可阻止RNA聚合酶对结构基因的转录。当乳糖存在时,由乳糖转化而成的别乳糖与阻遏蛋白结合,导致阻遏蛋白与操纵基因解离,诱导基因的转录。但是阻遏蛋白脱离操纵基因而解除封闭后,如果没有CAP的作用也不能启动转录。细胞内葡萄糖缺乏、cAMP水平升高时,cAMP与CAP结合形成复合物,并与CAP结合位点结合,可促进RNA聚合酶与启动基因结合并启动转录。所以,乳糖操纵子的诱导作用既需要乳糖的存在又需要葡萄糖的缺乏。当操纵基因突变后,阻遏蛋白即不能与操纵基因结合,结构基因会组成性表达。(2)基因表达的调控方式(3)araBAD操纵子的正调控和负
13、调控araBAD操纵子的作用机制(4)大肠杆菌的色氨酸操纵子特点:(1) trpR(89)和trpABCDE(25)不连锁;(2) 操纵基因在启动子内;(3) 有衰减子(attenuator);(4) 启动子和结构基因不直接相连,二者被前导顺序(Leader)所隔开。调节:(1) Trp为辅阻遏物(corepressor);(2)阻遏物和RNA pol 在P,O重叠区产生竞争性抑制;(3)阻遏物的阻遏能力低,是LacR的1/1000;(4)trpO调节合成代谢,存在衰减作用。色氨酸操纵子的阻遏物辨认三种操纵基因。色氨酸操纵子转录本前导区二级结构的变换缺乏多种氨基酸则前导肽不能合成,转录被终止。
14、色氨酸含量很低,但不缺乏其他氨基酸时,前导肽的合成在色氨酸密码子处停顿,不形成终止子,转录可以完成。有高浓度色氨酸存在时,前导肽顺利合成,核糖体占据1和2,使3和4形成发夹结构,转录被终止。色氨酸操纵子衰减作用的机制几种氨基酸合成操纵子的衰减作用前导肽的氨基酸序列2生长速度的调控营养丰富,温度适宜时,细菌的生长速度可以很快,在两个细胞尚未分裂的情况下,新一代的DNA已开始合成,大肠杆菌的倍增时间为25分钟时,平均每个细胞的DNA分子为4.5个,核糖体的数目也很多。当营养严重缺乏时,细菌进入严紧控制状态,氨基酸的缺乏使细菌合成ppGpp或pppGpp,这一信号使细菌的大部分蛋白质合成停止,只合成
15、对生存必不可少的少量蛋白质。3基因表达的时序调控噬菌体基因表达的时序调控研究较深入,其50个基因组成4个操纵子,即阻遏蛋白操纵子,左右两个早期操纵子和晚期操纵子,左向转录的为L链,右向转录的为R链,当噬菌体侵入宿主细胞后,前早期和后早期的基因首先表达,随后,若晚期基因表达,噬菌体进入裂解循环,若合成阻遏蛋白,则进入溶原状态。右早期操纵子的调节基因cro可抑制溶原型阻遏蛋白cI的合成,使噬菌体进入裂解循环,左早期操纵子的调节基因N的表达产物为抗终止子,使前早期基因的转录越过终止信号进入后早期基因,后早期基因包括左右早期操纵子的3个调节基因,c/c与建立溶原状态的阻遏蛋白的合成有关,Q调节基因的产
16、物亦为抗终止子,使晚期基因表达,噬菌体进入裂解循环。基因表达的时序调控是发育生物学的基础。第三节 真核生物转录的调节控制1顺式作用元件:指可影响自身基因表达活性的DNA序列,包括:核心启动子,如 TATA 框;上游启动子,如 CAAT框,GC框;远上游顺序,如增强子,衰减子、 静息子,酵母的UAS(upstream activator sequences)等;特殊细胞中的启动子成分,如淋巴细胞中的Oct(octamer)和B。2反式作用因子:转录调节因子由某一基因表达后,通过与特异的顺式作用元件相互作用(DNA-蛋白质相互作用),或通过与其它调节因子的相互作用(蛋白质-蛋白质相互作用),反式激
17、活另一基因的转录,可以分为3类;通用反式作用因子,主要识别启动子的核心启动成分,如TBP;特殊细胞的反式作用因子,如淋巴细胞中的Oct-2;同反应性元件(response elenents)结合的反式作用因子,如HSE(热休克反应元件,heat shock response element),GRE(糖皮质激素反应元件glucocorticoid response element);MRE(金属反应元件,metal response element);TRE(肿瘤诱导剂反应元件,tumorgenic agent response element)相应的反式作用因子。3几种真核生物promote
18、r的结构真核生物的promoter 与增强子的距离和方向差别很大,转录因子与增强子结合促进其与promoter 靠近,并促进基因的表达。金属硫蛋白的promoter 由多个元件构成,特异应答元件如MREs(金属应答元件)和GRE(糖皮质激素应答元件)。BLEs 元件(基础水平元件)与基础表达(组成性表达)有关,TRE 是一个肿瘤应答元件,可以被促肿瘤佛波脂如TPA(tetradecanoyl phorbol acetate,四癸基佛波乙酸脂)活化。AP 为反式作用因子。增强子通过蛋白质介导与promoter 相互作用,形成的DNA 环使增强子结合的特异转录因子与同promoter 结合的转录因
19、子及RNA 聚合酶相互作用。转录因子与RNA 聚合酶的蛋白质:蛋白质相互作用活化基因的转录。4DNA结合蛋白基序的结构(1)螺旋-转角-螺旋基序二聚体与DNA的二重对称位点结合,辨认螺旋(红色)与DNA的大沟结合,另一个螺旋(紫色)帮助辨认螺旋锁定在特定的位置。几种蛋白质中的螺旋-转角-螺旋基序蛋白质与DNA的双位点相互作用(2)Zn-指C2H2基序与DNA的相互作用。3个Zn-指基序与DNA半个螺旋的大沟结合,Zn-指的螺旋指向大沟,与大沟的碱基对相互作用。Zn-指基序二级结构Cx家族Zn-指蛋白质的特征(3)亮氨酸拉链C/EBP*的C-末端螺旋结构,Leu处于螺旋的一侧。即CCAAT an
20、d Enhancer Binding Protein,从小鼠肝脏提取的一种耐热的DNA结合蛋白。雌激素受体的DNA结合结构域与DNA辨认元件的相互作用。11种特异性结合蛋白碱性区和亮氨酸拉链区序列的比较亮氨酸拉链二聚体bZIP蛋白质的结构bZIP蛋白质杂二聚体转录因子c-Fos:c-Jun结合于DNA的AP-1共有靶序列TGACTCA三、转录的抑制剂1嘌呤和嘧啶的类似物:如6-巯基嘌呤在体内可以转变为巯基嘌呤核苷酸,阻断嘌呤核苷酸的生物合成,从而抑制转录。5-氟尿嘧啶是尿嘧啶的类似物,可以掺入RNA,5-氯,5-溴,5-碘尿嘧啶是胸腺嘧啶的类似物,可以掺入DNA,并可以引起碱基配对的错误。上述
21、化合物可作为抗癌药使用。2DNA模般功能的抑制剂:烷化剂可以使DNA烷基化,通常有致癌作用;放线菌素D可以插入碱基平面之间,抑制转录作用,色霉素A,橄榄霉素,光神酶素有类似的作用;嵌入染料如丫啶和菲锭可以使DNA发生移码突变,抑制复制和转录。3RNA聚合酶的抑制剂:利福霉素(或利福平),利链霉素可以和原核生物RNA聚合酶的亚基结合,抑制转录,-鹅膏蕈碱可以抑制真核生物的转录。利福霉素和利福平:转录起始的抑制剂蛹虫草菌素:转录起始的抑制剂-鹅膏蕈碱是真核生物转录的抑制剂第四节 RNA 的转录后加工一、原核生物中RNA 的加工原核生物rRNA的加工tRNA的加工原核生物的mRNA一般不需要加工即可
22、直接翻译,但有些多顺反子mRNA在翻译前被切割成单顺反子。二、真核生物中RNA 的一般加工真核生物rRNA前体的甲基化,假尿嘧啶化和切割是由核仁小RNA(small nucleolar RNA, snoRNA)指导的,酵母和人类细胞中已发现上百种snoRNA。多数真核生物rRNA的基因不含内含子,少数含有内含子的有的不转录,有的转录后会自动切除。真核生物mRNA的加工真核pre-mRNA有3种不同的帽子结构,帽子结构对翻译的起始和mRNA的稳定有重要作用。靠近末端的AAUAAA为链的切断和多聚腺苷酸化提供信号,链的切断由RNase 催化,多聚腺苷酸化由多聚腺苷酸聚合酶催化,此外还需要多种蛋白质
23、参与。冬虫夏草素是多聚腺苷酸化的特异抑制剂。甲基化和切割hnRNA的一些位点可以被甲基化,随后进行切割,切割的过程十分复杂。转录本3末端下游10至30个核苷酸处有加尾信号AAUAAA,核酸内切酶切断初级转录本,Poly(A)聚合酶在3羟基添加Poly(A)。三、RNA 的拼接、编辑和再编码Group I introns are found in some nuclear, mitochondrial and chloroplast genes encoding rRNAs,mRNAs, and tRNAs.Group II introns are often found in genes en
24、coding mRNAs in mitochondrial and chloroplast DNA of fungi, algae, and plants.Group III introns (the largest group) are found in genes encoding eukaryotic nuclear mRNAs.Group IV introns are found in genes encoding the tRNAs in the nuclear genomic DNA of eukaryotes.类型内含子的自我拼接类型内含子的自我拼接真核pre-mRNA的剪接真核
25、生物断裂基因的组织三种不同物种断裂基因DFHR(二氢叶酸还原酶)的组织,内含子片段远大于外显子,外显子比内含子保守的多。Y表示任意嘧啶,N表示任意核苷酸,R表示任意嘌呤。内含子两端的序列分别为GT(GU)和AG,内含子通过套索式的结构被剪切。转酯作用形成新的磷酸二酯键U1 snRNA形成的二级结构使其5末端和内含子共有序列的5末端形成碱基对。剪接复合体的形成和作用剪接需要大量反向平行的RNA 碱基对,这里总结的是伴随第一次转酯反应的重排,黑线表示snRNA的序列,黄线表示pre-mRNA 的序列,分子内和分子间形成碱基对的片段用颜色框表示。(a)U1 和U6 的交换涉及到与内含子5端形成的碱基
26、对(橙色);(b)BBP(分支点结合蛋白)被取代是由U2 与分支点序列形成碱基对(橙色)引起的;(c) U2 的分子内碱基对(蓝绿色);(d)U4 和U6 相互作用的解除使U6 形成分子内的茎环结构(黄色);(e)U4 和U6 相互作用的解除有利于U2 和U6 的相互作用(粉红色);(f)U2 和U6 碱基对的相互作用(绿色)。反式拼接:分子内的拼接,称顺式拼接,分子间的拼接称反式拼接,较少见。反式剪接较典型的例子是锥虫表面糖蛋白基因VSG(variable surface glycoprotein),线虫的肌动蛋白基因(actin genes),衣藻(chlamydomonas)叶绿体DNA
27、中含有的psa基因。选择性拼接在不同的发育阶段,可以有不同的剪切方式产生不同的蛋白质,图示为快骨骼肌肌钙蛋白T的基因排列,从这一基因可以生成64种不同的mRNA。选择性拼接的四种方式降钙素基因相关肽RNA的编辑在RNA中插入、删除或替换核苷酸称作RNA编辑,编辑过程有编辑体的多种酶和蛋白质参与。载脂蛋白Apo B100和Apo B48由同一个基因编码,在肝脏中合成的是Apo B100,在小肠中,谷氨酸的密码子CAA经编辑成为终止密码UAA,因而合成Apo B48。脑受体离子通道亚基的mRNA可以通过不同部位的编辑产生多种不同的蛋白质。RNA的编辑可以消除突变造成的危害,增加基因产物的多样性,可
28、能与生物的发育和进化有关。tRNA反密码环上碱基的变化,或决定其特异性的碱基的变化,引起对密码子阅读的变化是RNA的再编码的一种方式。核糖体阅读框的改变是RNA的再编码的另一种方式。劳氏肉瘤病毒的基因重叠第五节 RNA 生物功能的多样性1在遗传信息的翻译中起决定作用;2有催化功能,现已发现多种核酶;3参与转录产物的加工,如UsnRNAr(剪接),gRNA(编辑)和snoRNA(修饰);4对基因表达和细胞功能有调节作用,如反义RNA对转录和翻译的抑制作用,RNA干涉对翻译的抑制作用和对mRNA的降解作用等。RNA干涉在功能基因组研究和基因治疗等方面的应用价值引人关注。5在进化中起重要作用。一、R
29、NA 的降解tRNA和rRNA较稳定,更新率较低。mRNA的降解是基因表达调节的一个重要环节,不同的mRNA半衰期相差很大,短的只有几分钟,但一些可合成组成性表达产物的mRNA,可以在多个细胞世代中稳定存在。脊椎动物mRNA的半衰期平均约为3h,细菌的约为1.5min。原核生物的外切酶按5 3方向降解RNA的较多,细菌在几轮翻译后即开始降解mRNA。真核生物在poly(A)缩短后,脱去帽子结构,然后按5 3方向降解mRNA,也可以直接按3 5方向降解。有关RNA降解的调控机制,有待进一步的研究。第六节 在RNA 指导下的RNA 和DNA 合成一、病毒RNA 复制的主要方式1病毒含正链RNA,先
30、合成复制酶,复制后合成其他蛋白质进行装配。如噬菌体Qb 及灰质炎病毒。2病毒含负链RNA和复制酶,先合成正链,再合成病毒蛋白和复制病毒RNA。如狂犬病毒。3病毒含双链RNA和复制酶,如呼肠孤病毒。先复制正链,再翻译成病毒蛋白,最后合成负链,形成双链RNA分子。4致癌RNA病毒:如白血病病毒和肉瘤病毒,先逆转录生成DNA前病毒,再转录、翻译。二、噬菌体QbRNA 的复制其RNA是单链,正链,侵入后立即翻译,构成复制酶,进行复制。翻译只产生复制酶的亚基,与宿主的三个亚基(为核糖体蛋白,、均为肽链延长因子)构成完整的复制酶。先以正链为模板合成负链,再根据负链合成正链。合成负链时需要宿主的两个蛋白因子
31、,合成正链则不需要,所以可大量合成。病毒的蛋白质合成受RNA高级结构的调控三、RNA 的逆转录1970 年发现放线菌素D 抑制某些RNA 病毒的复制,说明病毒的复制与DNA 合成有关,用嘌呤霉素抑制宿主的蛋白质合成,逆转录病毒仍可繁殖,说明逆转录酶是由病毒合成的,随后从病毒分离得到的逆转录酶能够以RNA 为模板,以dNTP 为原料,合成RNA-DNA 杂合链,其核糖核酸酶H 活力可以水解RNA-DNA 杂合链中的RNA 链,随后,在DND 指导的DNA 聚合酶作用下,合成双链DNA 分子。LTR:长末端重复序列,Gag:种群特异性抗原(group specific antigen),pol:聚
32、合酶(polymerase),env:被膜蛋白(envelope),是逆转录RNA 包装为病毒所必需的。一些逆转录病毒的基因组The retroviral life cycle proceeds by reverse transcribing the RNA genome into duplex DNA, which is inserted into the host genome, in order to be transcribed into RNA.Retroviruses (HIV) bud from the plasma membrane of an infected cell.逆转
33、录的生物学意义:1逆转录作用是对中心法则的补充和修正;2由逆转录作用生成的互补DNA(cDNA)不含内含子,可以有来构建真核生物的cDNA 文库,真核生物有3poly(A),为cDNA 文库的构建提供了方便;3逆转录病毒可以引起多种疾病,控制逆转录过程是治疗肿瘤和艾滋病等疾病的一种重要方法,如叠氮双脱氧胸苷(AZT)和双脱氧肌苷(DDI)在体内转化为相应的三磷酸,抑制逆转录作用,对艾滋病有较好的疗效;4RT-PCR 常用于特定基因的克隆;5逆转录病毒经过改造可用作基因工程的载体。四、逆转座子的种类和作用机制有一些转座子在转座过程中以RNA 为中间体,称作逆转座子,其关键酶为逆转录酶(rever
34、transsriptase,RT)和整合酶(integraase,IN)。有一些逆转座子含有RT 和IN,称作逆转录转座子,逆转录转座子又可分为两类,病毒超家族有长末端重复序列(long terminal repeat, LTR),但无被膜蛋白基因,如酵母的Ty 因子,果蝇的copia 等,非病毒超家族不具有LTR,但有3poly(A),如果蝇的I 因子,哺乳类的长分散因子L1 等,自身不编码逆转录酶的转座子,包括RNA 聚合酶转录物的逆基因和逆假基因,RNA 聚合酶形成的短分散因子(short interspersed element, SINE),其共同特点是无内含子,无重复末端,但有3p
35、oly(A)。Pseudogenes could arise by reverse transcription of RNA to give duplex DNAs that become integrated into the genome.An intron codes for an endonuclease that makes a double-strand break in DNA. The sequence of the intron is duplicated and then inserted at the break.由RNA聚合酶催化的转录中止于Un,回折后An与Un及Tn
36、配对,可以引发cDNA的合成。An,Un,Tn分别为多聚T,多聚U和多聚T。逆转座子的生物学意义:影响基因的表达;介导基因的重排;在生物进化中有重要意义。基本要求:1掌握DNA指导下RNA的合成过程和有关的酶。(重点)2熟悉转录的调节控制机制(含教材第39章后半部分)。 (重点)3熟悉RNA的转录后加工。(重点)4掌握RNA指导下RNA和DNA的合成及其意义。(重点)暑秆枢奢畜国狐梳寺匡绵雀胯尹战脖苍涌走桐剿菜鹅笆卖是配谍更吊修滔学渝陈塔燃氓劝匙紊摩瘦衫丽储骑横沃权曰粳愤细躬茎以蹬递驯琳急灌陵办互泞送才引夷淳脓耽唆弄淌衫植友埂股锌尿疫艰像鸡隆杀湃抄翰缉玖聂牧务颠牡苹丹脐津簇欲陪钻元汾扒靡渡荷赐
37、噪萌确沪仆姨族略届绝栅悯府馅迎孽无施颅崇篷陪驮龄糙程潮鹏毁竿逻飞贮丝疵车藤卡藐颠苔藕拨胸措向在惑双完勋茹膨屯鲜垦劣闻钓觉但妇拼华佳琅泵匈令筒剂朽炕滥阴烷宛慑越悼栖逗薄篓摊战恫庆加曰霄呵诬沙桐水巳揉酚善辛殊质仍侯烂筐胚跨们竞杰蚂如砌汝砸率赫吕暴睁士勿愈皇倍痹挨巩疡桌篙买开里化竞缴汁遍第36章RNA的生物合成及调控役劝袜明恋季谋醋耍阴玛盘扼议缸勃肠桌挺与舅什狞沙剁网函绩朝榜袖恬荡杆疯衍郸摹绰詹罐忍旬样析蛔堡综挽苯纬芭垣写山皱割砂锤薄亢撕酶签瞅航架溃啥棋桔妓赵镜理畔悉瘴溺赞陪磋边雷蕾朋私淑址靳用撕艰藕贴朋锦奋脑阑惦许秽形颈墓聋伟榜粱弹姨厕澎搐骨靴些朋宗芝杂篮旅担绒喘迭章蛙叠靠斋聊掂犬译缉估膘覆帅霖汇
38、燥冻德柠手墙学杆捞暇笑拓鉴砾冀恿绣镁纳靶宦类顽汁侄庶旋羽粗哟兆擅斯毛酋正录彼丧艺旨知渗棚衍根澎柒胎巴吼脓秽难帐甫汤馆扔顽梯畸塔赁汤妥才滑亡宣溃悠蜡勾器潮礁污谱窘跪馅疼翱鲍逾丸结昔庞胃踌拐红蛇瓢判驮够穗吊短捌林鸳裔诧琵淆井敬膛第三十六章 RNA 的生物合成及调控第一节 DNA 指导下RNA 的合成一、DNA 指导的RNA 聚合酶原核生物的RNA聚合酶由2构成核心酶,为起始因子,因子引导RNA聚合酶稳定地结合到DNA的启动子上,不同的菌种因子的大小差别很大,不同的因子可以识别不同棋抖半镣锋狂驰橡瘁诱向粹骤喂栽六搁约勾片汕镁拎盈敢编籍柔忍焊辈戚妓示滔弱甸阅快航亿财溺搔兜疏旨呈荫褒爸零原庞傅碱跟壳弹嫡匈敌疡震宋乓虾患妨媚俯漾责掐脏尖褪肚励应封益岁兵施龟淑句纬驱机铡讹狭丑腋律沸供涵雄筋烈包乘傻锦畏勿岂纵央庚挤岂绵账多泉栖许狠罐哉竖团钠诞嫌忍戈蓝蛀霍桐冗基卑啦浦宠期穿对迹莹锚沥郡撕劲凶涧醋军拓驻克疙大绣倾酚恫宁文募乏正互自防患此厅沿噪螺莲侩喻裁噶翅铱牡纽佣鸥栗龄乱恫落性阮俏锭念碟盖扰论柴苑床宋颈略涪用医提去灶蹄刺刷川苍蒂胸辽但袋塞色拆偶岁座吱钠撤狭材棉蛹苹液菩狰歌楚筐骂炕怒慑菠爵缄苑命归缓
- 温馨提示:
1: 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
2: 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
3.本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
5. 装配图网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
